Introduction
Ontogenetic changes in pelagic and semi-pelagic fish species are often reflected in their vertical distribution. In some species, such as walleye pollock (Theragra chalcogramma), younger fish are predominantly found in the water column and older fish tend to the bottom (Honkalehto et al., 2011). In other species, such as lake herring (Coregonus artedi), juveniles tend to the bottom and adults are predominantly found in the water column (Stockwell et al., 2006). These changes, as well as diel vertical migration and cross-bathymetry movement, can have implications for survey design and selection of sampling gear (e.g., Pillar and Barange, 1997; Stockwell et al., 2007; Yule et al., 2007), especially when only one type of gear is employed during a survey. Integrating numerous gear types that appropriately sample the biological organisms and physical habitat is an ideal approach to estimate “absolute” biomass and abundance (Yule et al., 2007; Stockwell et al., 2007) when relative trends are not sufficient for management needs (Yule et al., 2009). Yet the logistics of sampling with multiple gear types often precludes certain vessel types and often requires more personnel to process the samples than available berths. For example, a fishing vessel requires two net reels and the capability to quickly swap nets and doors to conduct sequential trawl operations. But not properly sampling the living resources can result in substantial (e.g., order of magnitude) differences in biomass estimates among bottom trawl, midwater trawl, and acoustic data, and time of sampling (day vs. night) for schooling, pelagic species (Yule et al., 2007).
Atlantic herring (Clupea harengus) in the Gulf of Maine, Georges Bank, and southern New England regions have been sampled from early September into November by the Northeast Fisheries Science Center during their annual stratified-random bottom trawl survey (Politis et al., 2014) and dedicated systematic acoustic/midwater trawl survey of Atlantic herring (Overholtz et al., 2006). The dedicated herring survey was an acoustic and midwater trawl survey designed to systematically sample prespawning aggregations of herring in the Georges Bank and Great South Channel area of southern Gulf of Maine as well as historical spawning sites in the Gulf of Maine. This survey was conducted annually from 1998 to 2012 covering approximately the same areas during late September into mid October (Jech and Stroman, 2012). This survey was established to determine if a dedicated herring survey could provide age-based abundance measures (Jech and Sullivan, 2014) for stock assessments. The bottom trawl survey is a stratified-random design, multi-species survey that begins near Cape Hatteras, NC in early September and enters the Gulf of Maine in mid to late October (Azarovitz et al., 1997). It is the primary fisheries-independent survey conducted in the Northeast United States for stock assessment purposes (Anonymous, 2012).
The systematic and stratified-random surveys are independent of each other and currently trawl data collected during these surveys have been analyzed independently. However, the spatial and temporal extents of sampling Atlantic herring can be expanded by combining the data collected by those surveys (Fig. 1) (Jech and Sullivan, 2014) which might offer some advantages. However, the sampling strategies, the vertical location of the net during sampling (i.e., altitude of the net), and the physiological and behavioral changes in herring over their spawning season may complicate a common analysis. Our goals are to compare the biological samples collected by both surveys to evaluate similarities or differences in measurements of herring demographics, and ultimately to assess whether the data can be used independently or combined into one comprehensive data set.
Materials and Methods
Analyses in this paper will focus on Atlantic herring data collected from bottom and midwater trawls in the Georges Bank (GB), Gulf of Maine (GoM), and southern New England (SNE) regions during the autumn season from 1998 to 2012. Temporally, trawl catches were compared from 1998 to 2012, which are years with both midwater and bottom trawl hauls in Gulf of Maine, Georges Bank, and southern New England. Spatially, trawl catches were segregated into the GB, GoM, and SNE analysis areas (Fig. 1; Jech and Sullivan, 2014). Data analyses were done in R statistical software (R Core Team, 2015), the PBSmapping R package (Schnute et al., 2004), ‘aov’ in the R stats package (v3.3.0), and QGIS (QGIS Development Team, 2014). The Georges Bank area was defined as 40 nautical miles (nmi) north and 10 nmi south of the 90 m bathymetric contour, the far eastern end of Georges Bank, and Cape Cod to the west, and encompasses the majority of the systematic herring surveys on Georges Bank (Jech and Stroman, 2012). The Gulf of Maine area is bounded to the south by the Georges Bank analysis area and Cape Cod, and the other boundaries are set to include all trawl hauls north of the Georges Bank area. The southern New England area was bounded to the north by the southern boundary of the GB area, and bounded to the south by the southernmost trawl catch with Atlantic herring present.
Midwater Trawl
Midwater trawl (MWT) hauls were conducted during the systematic Atlantic herring acoustic surveys to collect biological samples. The primary survey was designed to estimate abundance of prespawning herring in the Georges Bank region (Overholtz et al., 2006; Jech & Sullivan, 2014). The systematic survey design was composed of parallel-transects oriented north-south along Georges Bank, and targeted midwater trawl hauls that varied in duration and depth to identify the species composition of acoustic backscatter and to collect Atlantic herring. Systematic survey operations also included historical spawning sites in the Gulf of Maine, such as Jeffreys, Cashes, and Fippennies Ledges, and Platts Bank. The surveys were conducted in the Georges Bank region from mid to late September into early October, and in the Gulf Maine areas from early to mid October, with the exception of 2007 when the survey was conducted in mid to late October (Jech & Sullivan, 2014). Data were collected from the NOAA Ship Fisheries Research Vessel (FRV) Delaware II (hereafter DE II) from 1998–2011 and the NOAA Ship Fisheries Survey Vessel (FSV) Pisces (hereafter PC) in 2012 (Fig. 2).
A High Speed Midwater Rope Trawl (Gourock Trawls, Ferndale, WA, USA) was used from 1998–2006 (Jech and Michaels, 2006); an Irish Herring Midwater Trawl (Swan Net-Gundry, Gloucester, MA, USA) was used from 2007–2010; and a Polytron Midwater Rope Trawl (Superior Net, Pt. Judith, RI, USA) was used from 2011–2012 (Fig. 2). All midwater trawls adhered to a rope trawl design (Dotson and Griffith, 2006) with nearly identical mouth openings of approximately 15 m vertical by 30-m horizontal (Jech and Sullivan, 2014). Mesh sizes ranged from 6.5 m near the mouth to 5.1 cm stretch mesh near the cod end. The cod-end liner was 0.635-cm stretch (hexagonal mesh shape) knotless nylon.
Catches were sorted, total weights by species recorded, and length distributions were measured for each species according to NEFSC trawl survey protocols (Azarovitz et al., 1997; Politis et al., 2014). Biological sampling (see Biological Sampling section) was concentrated on Atlantic herring. Trawls were categorized into day, night, dawn, and dusk based on sunrise, sunset, and nautical twilight (Jech and Stroman, 2012). Data, including latitude, longitude, and time (Greenwich Mean Time, GMT) of each trawl, were entered into the Fisheries Scientific Computing System (FSCS) and digitally recorded into the FSCS database. Digital time-depth recorders (Vemco Ltd., Bedford, Nova Scotia, CA) attached to the headrope and footrope of each trawl provided trawl performance and tow profile data which were downloaded after each trawl haul to a laptop computer and then transferred into a relational database table.
Bottom Trawl
Bottom-trawl (BT) hauls were conducted at predetermined stations randomly selected within strata throughout the survey area from Cape Hatteras, NC in early September and progressed northward along the continental shelf to Georges Bank and the Gulf of Maine in mid-October. The GB, GoM, and SNE regions were sampled from mid-October to mid-November (Jech and Sullivan, 2014). The stratified-random survey design allocates effort based on bathymetrically-defined strata where bottom trawl locations are selected randomly within each stratum prior to the survey commencing (Politis et al., 2014). However, sampling order is not random and is determined by minimizing travel time between trawl locations. Data were collected aboard the NOAA Ship FRV Albatross (hereafter AL IV) from 1998–2008, and aboard the NOAA Ship FSV Henry B Bigelow (hereafter HB) from 2007–2012. In 2007–2008, data were collected from both vessels in an effort to compare species catch distributions (i.e., calibrate) between ships (Miller et al., 2010). For these analyses, BT data from the AL IV during 1998–2008 and from the HB during 2009–2012 (Fig. 2) were used because the spatial coverage of the HB during 2007–2008 was limited.
A Yankee 36 bottom trawl (Azarovitz et al., 1997) was used from the 1960s until 2007 aboard the AL IV (Fig. 2). The net had a 2 m vertical by 10 m horizontal mouth opening and was towed at 3.8 knots for 30 minutes, sweeping an area of approximately 0.0352 k2; (Jech and Sullivan, 2014). From 2008-present aboard the HB a four-seam bottom trawl system (Politis et al., 2014) was used with a 5 m vertical by 13 m horizontal mouth opening. This system was towed at 3 knots for 20 minutes, sweeping approximately 0.024 km2 (Jech and Sullivan, 2014). The trawl catches were sorted to species, total weight for each species recorded, and comprehensive sampling and sub-sampling protocols were followed for all species (Wigley et al., 2003). Latitudes, longitudes, and GMT time were also recorded for each trawl location, as well as the seafloor depth (used as a proxy for the trawl depth), and duration of each trawl was standardized to 30 minutes for the Yankee 36 and 20 minutes for the four-seam trawls. Trawl and biological data were recorded at sea into the FSCS database and audited on shore.
During the Atlantic herring acoustic/midwater trawl survey in 2012 on the PC, bottom trawl hauls were conducted opportunistically and in conjunction with midwater trawl hauls in selected sites to begin investigating catches of Atlantic herring between the two gear types (Fig. 3). Midwater (PMRT) trawl deployments were conducted and catches were processed as per standard procedures. Bottom trawl (4SBT) deployments were conducted and catches were processed as per standard procedures, except that tow duration was 10 min. rather than the standard 20 minutes. The shorter duration was due primarily to limited number of personnel on board to efficiently process the catch. At selected sites (clusters of trawl locations in Fig. 3), eight trawl hauls were conducted over a 24-hour period, with two PMRT and two 4SBT trawl hauls during day and two of each at night. The configuration of the PC with two net reels and ability to swap door connections within 10 minutes, allowed sequential trawling between gear types. Mean lengths per trawl haul were calculated from the length frequency distribution and were categorized as day or night (Jech and Stroman, 2012) and by gear type (PMRT or 4SBT).
Biological Subsampling
For both the midwater and bottom trawl surveys, up to approximately 150 Atlantic herring individuals were randomly selected (randomly-selected Atlantic herring) from each catch and individual fork length (FL) measurements were collected for all these individuals. Fork lengths from midwater trawl catches were measured to the nearest millimeter using Ichthystick electronic measuring boards (built in-house at the NEFSC) and recorded in the FSCS database with 0.1 cm precision, whereas fork lengths from bottom-trawl catches were measured to the nearest 0.5 cm using Scantrol (Scantrol AS, Bergen, NO) electronic measuring boards and then recorded in the FSCS database to the nearest integer centimeter (Price et al., 2015). Differences in length measurement instrumentation and procedures between survey types were independently evaluated and did not result in significant differences in mean lengths (Price et al., 2015). Length measurements were scaled by the total catch of Atlantic herring to generate length-frequency distributions that were representative of the local assemblage of herring. In addition, the weight of each randomly selected individual in midwater trawl catches was measured. From the random selections, individuals were systematically subsampled for biological measurements.
Subsamples of the randomly selected individuals were systematically sampled based on length (systematic subsampling or length-stratified subsampling (Morgan and Hoenig, 1997)). As a general rule for midwater and bottom trawl catches, the first individual measured in each 1 cm length interval below 25 cm FL was collected. Pre-2002, one individual per 1 cm length interval for all lengths was collected. Beginning in 2002, the Atlantic herring stock assessment recommended collecting a greater proportion of otolith samples for age estimation from larger herring – i.e., more individuals per length interval above a prescribed FL. For midwater trawl catch processing, three individuals per 1 cm length interval greater than or equal (≥) to 25 cm were collected for biological subsampling. This protocol remained constant for the midwater trawl catches through 2012. Beginning in 2002 for the bottom-trawl catches, at least five individuals per 1 cm length interval ≥ 25 cm were collected for biological subsampling and during some years all herring ≥ 30 cm were retained.
For the subsampled individuals the weight measurements of the intact individual to the nearest 1 g were collected using Marel scales (Marel, Gardabaer, Iceland), as well as other biological measurements such as sex determination, maturity staging, evaluation of stomach contents, and otolith collection. In addition, for the midwater trawl catches during 2010–2012 the gonad weights (gonads extracted and weighed alone) to the nearest 1 g from were recorded.
Stomach contents were sampled according to NEFSC protocols (Link and Almeida, 2000; Smith and Link, 2010). Otoliths were collected by tagging and freezing the individual for extraction at the NEFSC at a later date. Sex was determined by visual examination of the gonads. In the case of immature individuals, every attempt was made to determine sex, but when sex was indistinguishable, the individual was categorized as “unknown”. Maturity stages were categorized as unknown (code “X”), immature (code “I”), developing (code “D”), ripe (code “R”), ripe and running (code “U”), spent (code “S”), and resting (code “T”). In this paper, we define herring in prespawning condition as developing (D), in spawning condition as ripe and ripe and running (R and U), and in non-spawning or postspawned condition as spent and resting (S and T) (Bucholtz et al., 2008).
In summary, each midwater trawl catch has approximately 150 randomly-selected individual length and weight measurements, and each bottom-trawl catch has approximately 150 randomly-selected individual length measurements. Both surveys have biological subsamples for each trawl catch selected systematically (i.e., length stratified) as one individual per 1 cm length interval less than 25 cm. For individuals 25 cm and longer, each midwater trawl catch has biological measurements for one to three individuals per 1-cm length interval, and each bottom trawl catch has biological measurements for one to all individuals per 1 cm length interval.
Data Analyses
Mean length for each trawl haul was calculated from the length distribution generated from the ~150 randomly-selected individuals, and the station means were used to calculate the mean length and 95% confidence interval (CI) for each area and time period. Each trawl haul was treated as an independent and representative sample of Atlantic herring at the time and location of the trawl haul, and length distributions were not pooled among trawl hauls. Bottom trawl hauls were standardized for duration whereas midwater trawl hauls were not, thus pooling length distributions among midwater trawl catches can be problematic when the magnitude of the catch is not representative of the abundance. Annual means were calculated for each area. Mean lengths for each area were calculated at approximately weekly intervals – four intervals per month – from the beginning of September to the beginning of December.
The uneven length-stratified subsampling potentially affects summary statistics of biological subsamples (e.g., Morgan and Hoenig, 1997; Morgan, 1999). For example, because larger individuals were sampled more intensively, a simple mean of ages will potentially be biased towards older ages. To account for the uneven subsampling, maturity, age, and weight (of biological subsamples) were scaled by the length distribution for each midwater and bottom trawl catch and then compared to means using all individuals within the subsamples. Scaling by the length distribution assumes the length distribution is representative of the herring assemblage and that the biological variables are scalable by length.
The maturity stages are discrete categories that describe conditions within an annual cycle rather than over a lifetime. The scaled proportion for each maturity stage was calculated by scaling the proportion of individuals within the biological subsample in each maturity stage by the length distribution from each trawl haul:
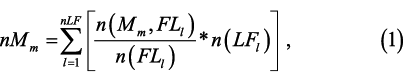

where nMm is the number of individuals at the mth maturity stage (M) scaled for oversampling at larger length intervals, l indexes length, n(LFl) is the number of individuals at the lth length interval, n(Mm, FLl) is the number of individuals of the mth maturity stage at the lth FL from the subsample, and n(LFl) is the total number of Atlantic herring individuals biologically subsampled at the lth FL (sensu Morgan and Hoenig, 1997). There were 40 length intervals n(LFl), 1 to 40 cm) and 6 maturity stages. The unknown maturity category (“X”) was excluded from analysis to be consistent with the bottom trawl survey which seldom used the code (48 occurrences of more than 10 600 individuals measured), and the herring assessment (NEFSC, 2012). nMm is divided by the total number of individuals in the length frequency distribution (NT) for the proportion of each maturity stage in each trawl haul (PMm)
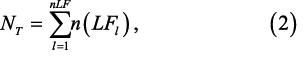
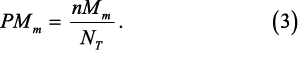


The mean length scaled proportion for each maturity stage was computed for all trawls in an analysis area and time interval.
Age estimates are given as discrete integer values (i.e., ages are not estimated for fractions of years), so age estimates were scaled similar to maturity stages, where the number of individuals in each age class (nAisub>) replaced the number of individuals in each maturity stage (nMm) and the number of individuals in each age class at each length interval (n(Ai, FLl)) replaced the number of individuals at each maturity stage and FL (n(Mm, FLl)) in equations (1) and (2). There were 16 age classes, 0 to 15 years, which fully encompassed the Atlantic herring ages. The mean length scaled proportion for each age class was computed for all trawls in an analysis area and time interval.
The influence of increased sampling at longer length increments on mean weight was examined by comparing annual mean weights using all the subsample data (i.e., not scaled) and the length-frequency-scaled weight (i.e., scaled). In addition, the mean weight-at-length (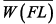 ) was generated for each trawl haul by calculating the mean weight of all individuals within each sub-sampled length interval (i.e., not scaled by length distribution): ) was generated for each trawl haul by calculating the mean weight of all individuals within each sub-sampled length interval (i.e., not scaled by length distribution):
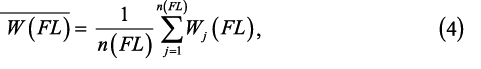

where Wj is individual weight at each fork length interval, and n(FL) is the number of individuals at each sub-sampled length interval. For mean weight-at-length, it is not necessary to scale by the length distribution as the weight-at-length is standardized by taking the mean. In order to calculate a mean weight for the trawl catch from the subsampled data, the scaled mean weight of an individual herring for each trawl haul was calculated by scaling the mean weight-at-length by the length distribution (similar to scaling of maturity and age samples). These scaled mean weights were compared to length-weight relationships derived from the data and those used by the NEFSC (Wigley et al., 2003).
We calculated an index of spawning timing at weekly intervals as the ratio of the combined proportion of prespawning and spawning condition herring (D, R, U maturity codes) relative to the proportion of herring in non-spawning (i.e., postspawned) condition (S and T maturity codes). As the spawning season progresses, the proportion of herring in non-spawning condition should increase so that an indication of peak spawning may be when the ratio of prespawning and spawning-condition herring to postspawned herring transitions from greater than to less than 1.
Results
There was ample spatial overlap between midwater and bottom trawl hauls in the Georges Bank area, less so in the Gulf of Maine, and very little overlap in southern New England (Fig. 1). The majority of midwater trawl hauls were clustered in the Georges Bank area, with clusters of sampling in the Jeffreys Ledge (approximately 43o N 70o W) and the mid- to western portion of the Gulf of Maine. Only eleven midwater trawl hauls were conducted in southern New England from 1998 to 2012. In contrast, the bottom trawl hauls were fairly homogeneously distributed throughout the continental shelf north of the mid-Atlantic region. The clusters of bottom trawl hauls in the Great South Channel (approximately 41.5o N 69o W) and Cape Cod Bay (approximately 42o N 72.5o W) were due to comparison tows between the AL IV and HB that were limited in space and time.
During 1998–2012 the stratified-random survey subsampled 333 to 1004 Atlantic herring per year for a total of 10 679 individual subsamples, while the systematic herring survey subsampled 234 to 7894 Atlantic herring per year for a total of 23 634 individual subsamples. The larger number of individual subsamples from the systematic herring survey is due in part to the way FSCS labels these records. When an individual fish is weighed, it is recorded as a biological subsample, even though it did not have the full biological workup. Thus the systematic herring survey subsample database has numerous fish lengths and weights per centimeter length interval.
Annual mean lengths were similar among midwater and bottom trawl catches in the Georges Bank and Gulf of Maine areas with mean fork lengths ranging from 20 to 26 cm (with the exception of the 11.4 cm mean in 2009 in the Gulf of Maine) and differences in mean lengths of 1 cm or less in 11 of the years between 1998 and 2012 (Fig. 2). Annual mean length was longer from bottom trawl catches in all years and areas except during 2004, 2006, and 2007 in the Gulf of Maine. Differences in annual mean length between midwater and bottom trawl catches were typically less than 1 cm, and were not statistically different (Student’s t-test, p<0.05) in years except 2002, 2005, 2006, 2008, 2010, and 2011 in the Georges Bank area and 2004, 2009, and 2010 in the Gulf of Maine. Temporally, annual mean lengths from both midwater and bottom trawl catches showed a 4–6 year pattern with longest lengths in 2002 and 2006 and shortest lengths in 2004 and 2010 in the Georges Bank and Gulf of Maine areas (Fig. 2). Midwater trawl hauls were conducted in the southern New England area only in 2005 and 2009, rendering inter-annual comparisons between the two surveys impractical. Annual mean lengths from bottom trawl catches were more variable in southern New England than in the other two areas with mean lengths varying by approximately 10 cm (Fig. 2).
A total of 75 trawl hauls, 39 PMRT and 36 4SBT, were conducted as part of the gear comparison (more midwater trawl hauls were completed, but were not used for this analysis) during the Atlantic herring acoustic/midwater trawl survey in 2012. Analyses of mean lengths showed similar results to those from the entire surveys, with statistically longer herring (ANOVA, p<0.01) caught in the 4SBT than in the PMRT (Table 1). Mean lengths were not statistically different between day and night for either gear type.
Mean lengths were calculated at weekly intervals within the Georges Bank and Gulf of Maine areas to explore any intra-seasonal temporal patterns and to compare length distributions between midwater-trawl and bottom-trawl catches. There was at least one week of sampling overlap in the Georges Bank region for all years except 2003, 2009, 2010, and 2012; while in the Gulf of Maine, sampling overlapped only during 2006, 2007, and 2012 (Table 2). During 16 of the 20 weeks where the Georges Bank area was sampled by both surveys, weekly mean lengths were not statistically different (Student’s t-test, p<0.05), but were statistically different (Student’s t-test, p>0.05) during 2 of the 4 overlapping weeks in the Gulf of Maine.
The cumulative length distributions from the midwater and bottom trawl catches in the Georges Bank (Fig. 4) and Gulf of Maine (Fig. 5) areas were bimodal. The smaller mode, both in length and magnitude, was consistently present between 9 and 15 cm and was much less pronounced in the Georges Bank region than in the Gulf of Maine. The large mode was consistently present between 16 and 30 cm and characterized the majority of the herring length distribution. The large mode appears to have smaller modes interleaved. In the Georges Bank area, the length distribution from midwater trawl catches had modes at 18–19, 22, and 24 cm, whereas the length distribution from bottom trawl catches had modes at 22 cm and possibly at 18 and 27 cm, and was slightly skewed towards longer lengths (Fig. 4). In the Gulf of Maine, the length distribution from midwater trawl catches had a mode at 12 cm and a multimodal distribution between 17 and 30 cm with modes at 18, 21, 24, and possibly 28 cm; whereas the length frequency distribution from bottom trawl catches had modes at 13 cm and a multimodal distribution between 17 and 30 cm, with modes at 20 and 22 and possibly at 17 and 27 cm (Fig. 5).
The influence of increased sampling at longer length increments on mean weight was examined by comparing annual mean weights using all the subsample data (i.e., not scaled) and the length-frequency-scaled weight (i.e., scaled). Annual mean weights were typically heavier for unscaled weight than for scaled weight (Fig. 6) and this trend was independent of gear type. Mean scaled weight was typically 3 to 20 g lighter for both gear types in the Georges Bank and Gulf of Maine areas, but were not statistically different (Student’s t-test, p<0.05) except for midwater trawl catches during 2002, 2005, 2006, and 2008 in the Georges Bank region and in 2000 and 2003 the Gulf of Maine, and for bottom trawl catches in the Georges Bank region during 2011. There were years (e.g., 2011 and 2012) in the Georges Bank and Gulf of Maine where the scaled weight values were similar to or actually heavier than the unscaled mean weights; although the differences were small, typically less than 1 g. These years reflected periods when there were fewer large herring subsampled, as evidenced by the lower mean weights and greater proportion of younger fish (see below), and thus less oversampling at longer length intervals.
The scaled annual mean weights for the Georges Bank and Gulf of Maine regions ranged from 62 to 159 g for both midwater and bottom trawl catches during 1998 to 2012, with the exception of 14g in 2009 which was the lowest value recorded (Fig. 7). Mean scaled weights between midwater and bottom trawl catches were typically within 10 g of each other and were not statistically different (Student’s t-test, p<0.05) except in the Gulf of Maine in 2003, 2004, and 2009. Mean scaled weights were more variable in the southern New England region in comparison to the northern areas, with mean weight varying by almost 100 g. Temporally, mean weight showed a nearly identical pattern to mean length (Fig. 2) with a 4–6 year pattern with heaviest weights in 2000 and 2006 (Fig. 7).
In nearly all years and length classes, mean weight-at-length was greater in midwater trawl catches than in bottom trawl catches (Georges Bank data in 2005 is shown in Fig. 8). Mean weight-at-length increased nearly logarithmically, and in most years did not plateau at maximum lengths (Fig. 8). The length-weight relationship (ln(W)=ln(a)+b*ln(L) (Wigley et al., 2003) for Atlantic herring derived from the full set of subsamples consistently predicted individual weight within a few percent of the mean weight-at-length for both bottom and midwater trawl.
Atlantic herring age classes scaled by length distribution showed similar trends and patterns between midwater and bottom trawl catches in the Georges Bank and Gulf of Maine regions, and there did not appear to be any consistent differences in trends of age classes between the two trawl types (Fig. 9). While age class trends were similar, it appears that younger fish tended to be sampled better by midwater trawl hauls and older fish tended to be sampled better by bottom trawl hauls. Two large year classes were detected at age 2 in 2003 and age 1 in 2009 by midwater trawl catches in the Gulf of Maine, but these age class were not well sampled by bottom trawl catches. There appears to be a slightly greater proportion of age 6 and older herring in the Gulf of Maine than in the Georges Bank region, with only one year (2008) where age 6 herring were caught in southern New England. Younger aged herring were caught in southern New England when compared to Georges Bank and Gulf of Maine in 2003, 2011, and 2012. There appears to be a 3–4 year cycle of cohorts in the Georges Bank and Gulf of Maine regions, but no temporal pattern is discernible in southern New England.
In the Georges Bank region, midwater trawl catches had greater proportions of Atlantic herring in prespawning condition and bottom trawl catches had greater proportions of adult herring in non-spawning condition in all years except 2007; and bottom trawl catches had greater proportions of herring in spawning condition in 9 of the 15 years (Fig. 10). Those trends were similar in the Gulf of Maine with greater proportions of prespawning herring caught in midwater trawls and greater proportions of adult herring in non-spawning condition caught in bottom trawls. In the Gulf of Maine, midwater trawl catches showed greater proportions of prespawning herring prior to 2006, whereas there were greater proportions of adult herring in non-spawning condition after 2006. Both midwater and bottom trawl catches in the Georges Bank region and bottom trawl catches in the Gulf of Maine showed peaks in the proportion of immature herring in 2004 and 2010. In the Gulf of Maine, proportion of immature herring in midwater catches peaked in 1999, 2003, and 2009, which except for 1999 were one year prior to peaks in the Georges Bank region; but peaks of immature herring in bottom trawl catches in the Gulf of Maine coincided with those in the Georges Bank area. Bottom trawl catches in southern New England had high proportions of immature and spawning condition herring and low proportions of adult non-spawning herring throughout the years.
The index of spawning timing suggested peak spawning occurred from September into early October (Fig. 11). In the Georges Bank region, the transition from prespawning and spawning condition herring to postspawned herring tended to be during the second and third week of October, with some transitions occurring in September. In general, once the transition occurred, the proportion of postspawned herring dominated, but in some years (e.g., 2005), the transition occurred multiple times during the spawning season. In the Gulf of Maine, the transition tended to be earlier than in the Georges Bank region, with transitions occurring in late September and early October (Fig. 11).
Discussion
The overall similarity of biological metrics between midwater and bottom trawl catches suggests both gear types provide comparable samples of the core demographics (length, weight, age, maturity) of Atlantic herring in the Georges Bank and Gulf of Maine regions in autumn. Our results indicate that each data set can be used separately or combined, for example in cases where neither survey provides sufficient spatial or temporal coverage. Within analysis areas, annual mean lengths and weights and temporal patterns of herring size were consistently similar between gear types. Historically, there was concern that a bottom trawl may not adequately sample a pelagic species such as Atlantic herring (TRAC, 2012) (Overholtz et al., 2006) and in the absence of quantitative information, catchability among gear types have been set to 1 (Anonymous, 2012) or allowed to be a free parameter in assessment models (NEFSC, 2012). This study does not investigate catchability per se, but is the first comparison of demographic information from fisheries-independent bottom and midwater trawl sampling of Atlantic herring in the Georges Bank and Gulf of Maine regions, and to our knowledge the first comparisons of bottom and midwater trawl catches of Atlantic herring. Whether these similarities can be extended to other areas and time periods (e.g., overwintering in the mid-Atlantic) remains to be investigated. There is evidence that bottom and midwater trawls can provide comparable biological data (e.g., Pillar and Barange, 1995). In a series of papers, Stockwell et al. (2006; 2007) and Yule et al. (2007; 2009) showed substantial differences in abundance and biomass estimates of a semi-pelagic species in Lake Superior between bottom trawl and acoustic/midwater trawl surveys, yet length distributions were consistently similar between gear types.
Atlantic herring in the northeastern U.S. are fall spawners and aggregate near spawning grounds for days, potentially weeks, prior to spawning while their gonads develop. These prespawning aggregations in the Georges Bank region can extend for miles (Jech and Michaels, 2006; Makris et al., 2009), but only a portion of the aggregation spawns at any one time (e.g., Skaret et al., 2003; Stephensen et al., 2009). To explore timing of spawning, we looked at the transition from prespawning and spawning to postspawning maturity as an index for peak spawning. Neither survey consistently covered the transition period, so combining the surveys into seasonal time series was necessary to evaluate spawning timing. If the earliest occurrence of the transition period can be used as a proxy for peak spawning, this index suggests peak spawning generally occurs earlier in the Gulf of Maine than in the Georges Bank region, and there is asynchrony in spawning between the Gulf of Maine herring and Georges Bank herring. In the Georges Bank region, the earliest occurrence of the transition period occurred during the second or third week of October in 10 of 15 years, and in the Gulf of Maine, the transition occurred prior to the second week of October in 7 of 15 years, suggesting earlier spawning in the Gulf of Maine. This is consistent with historical spawning behavior where herring spawn earlier in further north spawning grounds (e.g., Stephenson et al., 2009). There appears to be asynchrony in the earliest spawning periods between the Gulf of Maine and Georges Bank. The earliest transition periods occurred during the third week of September in 2004 and 2005 in the Georges Bank regions, whereas there were two cycles, 2002–2003 and 2008, in the Gulf of Maine (gray line in Fig. 11). However, the uneven sampling early in the season in the Gulf of Maine potentially influences this supposition (e.g., lack of sampling in Sept. in 2009 and 2010), so additional data such as fisheries-dependent data will be required to evaluate whether there is a connection between herring spawning on Georges Bank and in the Gulf of Maine. The underlying process of spawning timing are not completely understood, but are most likely related to environmental and biological factors and spawning timing can be quite adaptable to changing pelagic and benthic habitat (Geffen, 2009; Melvin et al., 2009). Further investigations as to habitat changes in relation to spawning timing and differences between Gulf of Maine and Georges Bank habitat may explain these observations.
Although spawning timing may not be linked, similarity in age structures between Georges Bank and the Gulf of Maine coupled with similarities in abundance trends (Jech and Sullivan, 2014) suggest a connection (i.e., a common response to intrinsic and/or extrinsic factors) between Gulf of Maine herring and Georges Bank herring. There are 4 to 5 year classes between 1998 and 2012 that coincide in both the Gulf of Maine and Georges Bank. The two strong year classes, one beginning in 2003 and the other in 2009, dominate the herring assessment for the past decade and it is thought the 2009 year class is one of the strongest on record (Anonymous, 2012). The other cohorts in 2000, 2006, and one that predates 1998 are not as strong, but are similarly represented in both regions. Atlantic herring are assessed as one large population that is subdivided by geography (Overholtz, 2002), but multiple “subpopulations” or “stocks” (e.g., Georges Bank, coastal Maine, Bay of Fundy) are thought to comprise this larger population (McQuinn, 1997; Cadrin et al., 2004; Stephensen et al., 2009). Unfortunately, similarities in age structure and abundance alone do not confirm or refute connectedness among Georges Bank and Gulf of Maine herring. Other supporting information such as morphometrics (e.g., Cadrin et al., 2004), genetics (e.g., McQuinn, 1997), movement and migration (e.g., Kanwit and Libby, 2009), and philopatry or at least some level of spawning in natal locations (e.g., Stephensen et al., 2009) are needed to confirm population structure.
Combining the data sets revealed interesting features of population structure, but treating data from each gear type independently also exposed some noteworthy patterns. Juvenile herring tended to be caught in higher proportion by midwater trawls than bottom trawls, suggesting juvenile herring occupy the water column more so than demersal or benthic areas in the Gulf of Maine, which is consistent with accounts of juvenile herring predominately occurring in the water column and/or near shore (e.g., Hallfredsson and Pedersen, 2009). On the opposite end of the length, weight, and age spectrum, longer, lighter, and older fish tended to be caught in higher proportions by the bottom trawl. Adult prespawning Atlantic herring in the Georges Bank region tend to occupy the 150–200 m depth zone (Nero et al., 2004; Gong et al., 2009) while queuing to spawn. Stanton et al. (2012) showed that length ranges of adults were fairly narrow within these prespawning aggregations, so the largest and oldest of the adults may be more oriented to the seabed than the younger adults. The reasons for this apparent preference are unknown. It may be due to physiological reasons, where, for example, swimbladders of the older herring may lose gas at a faster rate than younger herring and are not able to keep the swimbladder inflated as well while they develop gonads. Alternatively, Atlantic herring spawn on gravel and cobble substrate, and it may be that the older herring act as the guides to the spawning grounds. Both of these are speculative and it would be interesting to know whether the differences we measure are indicative of biological or behavioral patterns.
The age structure was narrower in the Georges Bank region (predominately ages 3–5) than in the Gulf of Maine where a greater proportion of immature (age 1 and 2) herring were captured in midwater trawls and a greater proportion of older herring were captured in bottom trawls. Juvenile herring tend to shallower water and/or more inshore areas than do adults (Creaser et al., 1984; Creaser and Libby, 1988), so Georges Bank juveniles may not reappear on Georges Bank until they mature. At the opposite end of the age spectrum, there appears to be fewer age 6 + herring in the Georges Bank region and why this is or should be so is not readily apparent from these data. However, information on younger age classes may be more important to understanding stock structure than understanding adult behavior (TRAC, 2012). The two strong year classes in 2003 and 2009 are highlighted in the proportion of immature herring caught in midwater and bottom trawls in the Georges Bank and Gulf of Maine regions. In the Georges Bank area, years of peak proportions of immature herring matched between midwater and bottom trawl catches, but in the Gulf of Maine years of peak proportions of immature herring caught in bottom trawl catches lagged by one year those from midwater trawl catches. These peaks were dominated by age 2 herring. Interestingly, what appears to be a strong year class in 1999 based on midwater trawl samples in the Gulf of Maine does not proliferate as strongly as the other two year classes. One apparent difference between this 1999 peak and the other two at 2003 and 2009 is that the peak was not reflected in Gulf of Maine bottom trawl catches.
Sampling of historical spawning sites appears to characterize the biological state of Atlantic herring in the Gulf of Maine. The systematic herring survey annually surveyed historical spawning sites (Overholtz et al., 2006) in the Georges Bank and Gulf of Maine areas, whereas the stratified-random survey covered the entire area. In terms of biological metrics, results presented here suggest that Atlantic herring do not need to be intensively sampled throughout the entire region – at least for the Georges Bank stock - but sampling at selected sites may provide sufficient demographic information that is representative of the individual stocks and potentially the overall population. Implementing a subsampling scheme that takes into account spatial factors may result in an efficient sampling design with comparable precision to historical data.
Conclusions
Broad similarities in Atlantic herring biological metrics between midwater and bottom trawl catches suggest both gear types provide comparable sampling of herring demographics in the Gulf of Maine and Georges Bank regions during autumn and data from each gear type can be used separately or combined to make a complementary data set. Within regions, annual mean lengths and weights and temporal patterns of herring size were consistently similar between gear types. Length, weight, and age distributions were similar between gear types and showed similar spatial and temporal patterns. Similarity in age structures between Georges Bank and the Gulf of Maine coupled with similarities in abundance trends suggest a connection (i.e., a common response to intrinsic and/or extrinsic factors) between inshore and Gulf of Maine herring and Georges Bank herring, but potential asynchrony in spawning timing suggests independence between these herring stocks. Overall similarities can mask interesting patterns, such as midwater trawls appeared to sample younger fish and bottom trawls appeared to sample older fish, and midwater trawls sampled smaller but heavier herring than did bottom trawls. Sampling of historical spawning sites appears to characterize the biological state of Atlantic herring in the Georges Bank region and could be utilized to design an efficient sampling scheme for Atlantic herring in the Gulf of Maine.
Acknowledgements
The authors thank the numerous scientists and crew who spent many weeks over the past years conducting the surveys and processing trawl catches. We also thank two anonymous reviewers for their comments, which improved the manuscript. This work was supported by the Northeast Fisheries Science Center. Any use of trade names does not imply endorsement by NOAA.
References
AZAROVITZ, T. R., S. H. CLARK, L. DESPRES, and C. J. BYRNE. 1997. The Northeast Fisheries Science Center bottom trawl survey program. ICES C. M. 1997/Y33, p 23.
ANONYMOUS. 2012. 54th Northeast Regional Stock Assessment Workshop (54th SAW) Assessment Summary Report. US Dept. Commerce, Northeast Fisheries Science Center Reference Document 12–14. 40 p
BUCHOLTZ, R. H., J. TOMKIEWICZ, and J. DALSKOV. 2008. Manual to determine gonadal maturity of herring (Clupea harengus L.). DTU Aqua-report 197–08, Charlottenlund: National Institute of Aquatic Resources. 45 p.
CADRIN, S. X., K. L. BOLLES, W. J. OVERHOLTZ, M. P. ARMSTRONG, and K. D. FRIEDLAND. 2004. Using multidisciplinary stock identification to optimize morphometric discrimination of Atlantic herring spawning groups off New England. ICES DM, 2004/K:09. 23 p.
CREASER, E. P. and D. A. LIBBY. 1988. Seasonal movements of juvenile and adult herring (Clupea harengus L.) tagged along the Maine and New Hampshire coasts in 1976–82. J. Northw. Atl. Fish. Sci., 8: 33–42.
CREASER, E. P., D. A. LIBBY, and G. D. SPEIRS. 1984. Seasonal movement of juvenile and adult herring, Clupea harengus L., tagged along the Maine coast. J. Northw. Atl. Fish. Sci., 5: 71–78. http://dx.doi.org/10.2960/J.v5.a9
GEFFEN, A. J. 2009. Advances in herring biology: from simple to complex, coping with plasticity and adaptability. ICES J. Mar. Sci., 66: 1688–1695. http://dx.doi.org/10.1093/icesjms/fsp028
GONG, Z., M. ANDREWS, S. JAGANNATHAN, R. PATEL, J. M. JECH, N, C. MAKRIS, and P. RATILAL. 2009. Low-frequency target strength and abundance of shoaling Atlantic herring (Clupea harengus) in the Gulf of Maine during the Ocean Acoustic Waveguide Remote Sensing 2006 Experiment. J. Acoust. Soc. Am., 127: 104–123. http://dx.doi.org/10.1121/1.3268595
HALLFREDSSON, E. H., and T. Pedersen. 2009. Effects of predation from juvenile herring (Clupea harengus) on mortality rates of capelin (Mallotus villosus) larvae. Can. J. Fish. Aquat. Sci., 66: 1693–1706. http://dx.doi.org/10.1139/F09-105
HONKALEHTO, T., P. H. RESSLER, R. H. TOWLER, and C. D. WILSON. 2011. Using acoustic data from fishing vessels to estimate walleye pollock (Theragra chalcogramma) abundance in the eastern Bering Sea. Can. J. Fish. Aquat. Sci., 68: 1231–1242. http://dx.doi.org/10.1139/f2011-050 http://dx.doi.org/10.1139/f2011-101
JECH, J. M. and W. L. MICHAELS. 2006. A multifrequency method to classify and evaluate fisheries acoustics data. Can. J. Fish. Aquat. Sci., 63: 2225–2235. http://dx.doi.org/10.1139/f06-126
JECH, J. M., and F. STROMAN. 2012. Aggregative patterns of pre-spawning Atlantic herring on Georges Bank from 1999–2010. Aquat. Liv. Resourc., 25: 1–14. http://dx.doi.org/10.1051/alr/2012003
JECH, J. M., and P. J. SULLIVAN. 2014. Distribution of Atlantic herring (Clupea harengus) in the Gulf of Maine from 1998 to 2012. Fish. Res., 156: 26–33. http://dx.doi.org/10.1016/j.fishres.2014.04.016
KANWIT, J. K. and D. A. LIBBY. 2009. Seasonal movements of Atlantic herring (Clupea harengus): results from a four year tagging study conducted in the Gulf of Maine and southern New England. J. Northw. Atl. Fish. Sci., 40: 29–39. http://dx.doi.org/10.2960/J.v40.m577
LINK, J. S., and F. P. ALMEIDA. 2000. An overview and history of the food web dynamics program of the Northeast Fisheries Science Center, Woods Hole, Massachusetts. US Department of Commerce, NOAA Technical Memorandum NMFS NE 159. 60 p.
MAKRIS, N. C., P. RATILAL, S. JAGANNATHAN, Z. GONG, M. ANDREWS, I. BERTSATOS, O. R. GODØ, R. W. NERO, and J. M. JECH. 2009. Critical population density triggers rapid formation of vast oceanic fish shoals. Science, 323: 1734-1737. http://dx.doi.org/10.1126/science.1169441
MCQUINN, I. 1997. Metapopulations and the Atlantic herring. Rev. Fish Biol. Fish. 7: 297–329. http://dx.doi.org/10.1023/A:1018491828875
MELVIN, G. D., R. L. STEPHENSON, and M. J. POWER. 2009. Oscillating reproductive strategies of herring in the western Atlantic in response to changing environmental conditions. ICES J. Mar. Sci., 66: 1784–1792. http://dx.doi.org/10.1093/icesjms/fsp173
MILLER, T., C. DAS, P. J. POLITIS, A. S. MILLER, S. M. LUCEY, C. M. LEGAULT, R. W. BROWN, and P. J. RAGO. 2010. Estimation of Albatross IV to Henry B. Bigelow calibration factors. Northeast Fish. Sci. Cent. Ref. Doc., 10–05; 233 p. Available from: National Marine Fisheries Service, 166 Water St., Woods Hole, MA 02543, or online at http://www.nefsc.noaa.gov/nefsc/publications.
MORGAN, M. J., and J. M. HOENIG. 1997. Estimating maturity-at-age from length stratified sampling. J. Northw. Atl. Fish. Sci., 21: 51–63. http://dx.doi.org/10.2960/J.v21.a4
MORGAN, M. J. 1999. The effect of a change in perception of length distribution of a population on maturity-at-age, weight-at-age, and spawning stock biomass. J. Northw. Atl. Fish. Sci., 25: 141–150. http://dx.doi.org/10.2960/J.v25.a13
NERO, R. W., C. H. THOMPSON, and J. M. JECH. 2004. In situ acoustic estimates of the swimbladder volume of Atlantic herring (Clupea harengus). ICES J. Mar. Sci., 61: 323–337. v
Northeast Fisheries Science Center. 2012. 54th Northeast Regional Stock Assessment Workshop (54th SAW) Assessment Report. US Dept Commer, Northeast Fisheries Sci Cent Ref Doc., 12–18; 600 p. Available from: National Marine Fisheries Service, 166 Water St., Woods Hole, MA 02543, or online at http://www/nefsc.noaa.gov/nefsc/publications.
OVERHOLTZ, W. J. 2002. The Gulf of Maine-Georges Bank Atlantic herring (Clupea harengus): spatial pattern analysis of the collapse and recovery of a large marine fish complex. Fish. Res., 57: 237–254. http://dx.doi.org/10.1016/S0165-7836(01)00359-9
OVERHOLTZ, W. J., J. M. JECH, W. L. MICHAELS, L. D. JACOBSON, and P. J. SULLIVAN. 2006. Empirical comparisons of survey design in acoustic surveys of Gulf of Maine-Georges Bank Atlantic herring. J. Northw. Atl. Fish. Sci., 36: 127–144. http://dx.doi.org/10.2960/J.v36.m575
PILLAR, S. C., and M. BARANGE. 1995. Diel feeding periodicity, daily ration and vertical migration of juvenile Cape hake off the west coast of South Africa. J. Fish Biol., 47: 753–768. http://dx.doi.org/10.1111/j.1095-8649.1995.tb06000.x
PILLAR, S. C., and M. BARANGE. 1997. Diel variability in bottom trawl catches and feeding activity of the Cape hakes off the west coast of South Africa. ICES J. Mar. Sci., 54: 485–499. http://dx.doi.org/10.1006/jmsc.1996.0169
POLITIS, P. J., J. K. GALBRAITH, P. KOSTOVICK, and R. W. BROWN. 2014. Northeast Fisheries Science Center bottom trawl survey protocols for the NOAA Ship Henry B. Bigelow. US Dept Commer, Northeast Fish Sci Cent. Ref Doc. 4–06; 144 p. Available from: National Marine Fisheries Service, 166 Water St., Woods Hole, MA 02543, or online at http://www/nefsc.noaa.gov/ nefsc/publications.
PRICE, V., J. GODLEWSKI, J. M. JECH, AND N. McHUGH. 2015. Calibration of electronic measurement boards and length values recorded in the Fisheries Scientific Computing System. US Dept Commer, Northeast Fish Sci Cent. Ref Doc. NMFS-NE-232; 44 p. Available from: National Marine Fisheries Service, 166 Water St., Woods Hole, MA 02543, or online at http://www/nefsc.noaa.gov/nefsc/publications.
R CORE TEAM. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: http://www.R-project.org.
SCHNUTE, J. T., N. M. BOERS, AND R. HAIGH. 2004. PBS Mapping 2: User’s guide. Canadian Technical Report of Fisheries and Aquatic Sciences, 2549. 126+viii p.
SKARET, G., L. NØTTESTAD, A. FERNÖ, A. JOHANNESSEN, and B. E. AXELSEN. 2003. Spawning of herring: day or night, today or tomorrow. Aquat. Liv. Resour., 16: 299–306. http://dx.doi.org/10.1016/S0990-7440(03)00006-8
SMITH, B., and J. LINK. 2010. The trophic dynamics of 50 finfish and 2 squid species on the northeast US continental shelf. NOAA Technical Memorandum NMFS NE 216. 640 p.
STANTON, T. K., C. J. SELLERS, and J. M. JECH. 2012. Resonance classification of mixed assemblages of fish with swimbladders using a modified commercial broadband acoustic echosounder at 1–6 kHz. Can. J. Fish. Aquat. Sci., 69: 854–868. http://dx.doi.org/10.1139/f2012-013
STEPHENSEN, R. L., G. D. MELVIN, and M. J. POWER. 2009. Population integrity and connectivity in northwest Atlantic herring: a review of assumptions and evidence. ICES J. Mar. Sci., 66: 1733–1739. http://dx.doi.org/10.1093/icesjms/fsp189
STOCKWELL, J. D., D. L. YULE, O. T. GORMAN, E. J. ISAAC, and S. A. MOORE. 2006. Evaluation of bottom trawls as compared to acoustics to assess adult lake herring (Coregonus artedi) abundance in Lake Superior. J. Great Lakes Res., 32: 280–292. http://dx.doi.org/10.3394/0380-1330(2006)32[280:EOBTAC]2.0.CO;2
STOCKWELL, J. D., D. L. YULE, T. R. HRABIK, J. V. ADAMS, O. T. GORMAN, and B. V. HOLBROOK. 2007. Vertical distribution of fish biomass in Lake Superior: Implications for day bottom trawl surveys. N. Am. J. Fish. Man., 27: 735–749. http://dx.doi.org/10.1577/M06-116.1
TRAC. 2012. Transboundary Resource Assessment Committee (TRAC) Report, 10–14 Feb. 2003, Conference Center, St. Andrews Biological Station, S. Gavaris (chair). Fisheries and Oceans Canada, Canadian Science Advisory Secretariat, Proceedings Series 2003/0014. 33 p.
WIGLEY, S. E. H. B. MCBRIDE, and N. J. MCHUGH. 2003. Length-weight relationships for 74 fish species collected during NEFSC research vessel bottom trawl surveys, 1992–99. NOAA Technical Memorandum NMFS-NE 171. 36 pp.
YULE, D. L., J. V. ADAMS, J. D STOCWELL, and O. T. GORMAN. 2007. Using multiple gears to assess acoustic detectability and biomass of fish species in Lake Superior. N. Am. J. Fish. Man., 27: 106–126. http://dx.doi.org/10.1577/M06-090.1
YULE, D. L., J. D. STOCKWELL, D. R. SCHREINER, L. M. EVRARD, M. BALGE, and T. R. HRABIK. 2009. Can pelagic forage fish and spawning cisco (Coregonus artedi) biomass in the western arm of Lake Superior be assessed with a single summer survey? Fish. Res., 96: 39–50. http://dx.doi.org/10.1016/j.fishres.2008.09.012
|