Introduction
The use of non-trawl gears in deepwater fisheries conducted in various areas around the world was documented in a review of the commercial potential of deepwater resources off Atlantic Canada commissioned by the Canadian Department of Fisheries and Oceans (DFO) Maritimes Region. The review report (Pohle et al., 1992) suggested that inclusion of fixed gears in exploratory fishing programs in Canadian waters, for both fishes and invertebrates, was worthy of consideration. In response, DFO conducted fishing trials using a variety of fixed gears on the Scotian Shelf slope, south of Nova Scotia, in 500–2800 m in August 1991. The present paper reports on the fish catches made during these trials.
The primary purpose of the venture was developmental – to examine the practicalities of fishing fixed gears of various types and sizes in deep water and to determine their success in catching fish and crustaceans. However, the results are also of scientific interest as fishing was conducted, in large degree, according to a sampling design that was stratified by depth and area of fishing. This allows an account to be given of the composition of the fish fauna on the continental slope as portrayed by fixed gears, and for comparisons to be made with catches from otter trawl surveys conducted at similar depths.
Methods
Gear trials were conducted from the DFO research vessel Alfred Needler, a 50 m stern otter trawler, during 7–12 August 1991 (cruise designation - N156). Gears deployed were primarily bottom gillnets but included crab and shrimp traps and bottom longlines.
Gillnets were of nylon; the smallest used (6.0–8.9 cm mesh sizes, multifilament twine) were typical herring and mackerel fishing nets, the midsized nets (14.0–17.8 cm mesh sizes, monofilament twine) were those used for groundfish fishing and the largest (26.7 cm mesh size) was a lumpfish (Cyclopterus lumpus) net. Table 1 gives net specifications and the number of times each was fished. Mesh sizes were rounded to the nearest cm for data presentation and the 14.0 and 15.2 cm mesh data were amalgamated and presented as being for 15 cm mesh nets. This amalgamation was justified by the observation that the numbers caught of each species were very similar for these mesh sizes and so too were the average lengths and weights of the most commonly caught species (Table 2). Centroscymnus coelolepis was an exception with larger sizes apparently being caught in 15.2 cm nets. However, this appeared attributable to an imbalance in the number of nets of each size fished at 1350 m in the Gully where particularly large specimens of this species were caught (see results).
Crab traps had a 100 kg conical steel frame (base diameter about 2 m) covered by 75 mm mesh polyethylene netting and equipped with four plastic entrance cones on top with minimum openings of about 35 cm. Shrimp traps were of steel mesh construction (1.27 cm bar length), rectangular in shape (about 75 cm long, 60 cm wide and 30 cm high) and equipped with entrances (of 2.5 or 5 cm minimum width) located either on top or on the side. Traps were baited with herring, squid or both, sometimes supplemented with other species. Twenty-three crab traps and 33 shrimp traps were set in total.
Longlines were deployed only once due to technical difficulties. The string consisted of Mustad circle hooks of sizes 10/0 (100 hooks), 12/0 (100 hooks) and 14/0 (200 hooks) baited with squid.
Gears were usually set in arrays consisting of either four gillnets or of two crab traps along with up to five shrimp traps. When setting gillnets, the sequence was to attach an anchor, consisting of 45 kg of chain, to a 180 m rope, the other end of which was attached to one end of the net array. The other end of the array was attached, by a 180 m or 360 m rope to another chain anchor, weighing 45–90 kg. From this anchor, a vertical rope extended to a buoy at the surface. The nets were attached to a headline consisting of 16 mm diameter poly-rope and to a ground line of 10 mm diameter No. 33 lead rope. Traps were set about 30 m apart along the ground line. Because of their weight, no anchors were considered to be necessary. Indeed, for several tows, a mixed array of gillnets and traps was deployed, the traps being set at either end to serve as anchors.
The setting and hauling of gear did not follow a fixed schedule but setting occurred most commonly in the afternoon or evening. For both gillnets and traps, soak times ranged from 11 to 43 hours but were 22–25 hours in half the cases and the modal value was 24 hours. The effect of soak time on catches is unpredictable and may be negligible (Bérubé et al., 2000). Thus, variations in soak time are ignored in the present analysis.

Fig. 1. Areas fished by the Alfred Needler in August 1991.
|
Sampling occurred in two areas, one south of Emerald Bank and one in The Gully (Fig. 1). The 15 cm gillnets and the traps were set in both areas but the other sizes of gillnet were set only in The Gully and longlines only off Emerald Bank. In each area, gears were set at depths of approximately 500, 900, 1350, 1800 and 2800 m. In total, 21 gear arrays were set but in three cases the gear was lost.
Water temperature data were collected using expendable bathythermographs. Three casts were made at each of the sampling areas, over bottom depths of about 400, 700 and 1700 m (about the maximum range of the instruments used), to provide estimates of near bottom temperatures at the depths fished.
Results
Water temperature
Temperatures near bottom at 500 m, the shallowest depth fished, were about 4.9°C in The Gully and 5.3°C off Emerald Bank. Temperatures converged with increasing depth and at 1 700 m, were about 3.5°C in both areas. Thus, at the same depths, temperatures in The Gully were slightly lower than south of Emerald Bank to a depth of 1 700 m, but likely the same at deeper depths.
Catches by all gears
Catches were comprised of about 1 400 specimens weighing 3.1 metric tons. Scientific and common names of the 21 taxa represented in these catches, the number and weight caught of each, and the depth range within which each was caught, are given in Table 3. None of the taxa represented new records for the area. Centroscyllium fabricii was dominant in catches numerically, comprising 43%. In terms of weight, however, C. coelolepis contributed about the same amount as C. fabricii and Hydrolagus affinis contributed more. Collectively, these three species accounted for 63% of catches by number and 73% by weight. The great majority of catches were taken in gillnets (87% by no. and 96% by wt.).
Catches by 15 cm mesh gillnets
The 15 cm mesh gillnets were the most extensively deployed. Four nets were fished at 900, 1350 and 1800 m in each sampling area, except that, at 1350 m in The Gully, seven nets were fished. Thus, for presentation, the data from The Gully/1350 m depth stratum were adjusted by x0.57 to standardize to a four-net unit. In addition, an array of 3 nets was fished at 2 800 m in The Gully but only three fish were caught in those, one each of H. affinis, Macrourus berglax and Antimora rostrata, suggesting a gear malfunction and these results are not considered comparable to those at shallower depths.
The number of fish caught in 15 cm gillnets in The Gully was 25% fewer than off Emerald Bank for the same fishing effort (12 nets set per area), but the weight of fish caught was 10% greater (Table 4). Centroscymnus coelolepis, C. fabricii and Etmopterus princeps were all much less numerous in The Gully catches and increased abundance of H.affinis and several other species in The Gully did not fully compensate, numerically, for these reductions. However, average weights of the main species caught were greater in catches from The Gully than in catches from off Emerald Bank (Table 2). Centroscymnus coelolepis was particularly influential, being 30% longer and three times heavier in Gully catches.
Total numbers caught per unit of effort (i.e. per eight nets set) decreased with depth, at 1800 m being only 40% of that at 900 m, but weight caught increased with depth, at 1800 m being 40% higher than that at 900 m. Thus, the average weight of an individual fish caught increased with depth, from 1.5 kg at 900 m to 5 kg at 1800 m (Table 4). This difference by depth in the ratio of overall weight to numbers caught was attributable largely to replacement of the relatively small-bodied species C. fabricii and E. princeps that dominated the 900 m zone with lesser numbers of the much larger H. affinis at greater depths (Table 4). Also, in some species, mean weight increased with depth, e.g. C. coelolepis and Reinhardtius hippoglossoides, but did not in others, M. berglax and A. rostrata.
Length frequencies of the six most commonly caught species (Fig. 2) show that few fish smaller than 40 cm were caught. Specimens of the largest species in catches, H. affinis, were 80–142 cm long.
Catches by gillnets with other mesh sizes
Small numbers of nets with mesh sizes of 6–9, 18 and 27 cm were fished in 900–1 350 m in The Gully and their catches are compared to those of the 15 cm mesh nets at 900 m at this location (Table 5). No attempt is made to standardize for the different sizes of nets but it can be concluded that the 18 cm mesh nets did not differ greatly from the 15cm nets in species, or numbers, caught whereas the substantially larger 27 cm net caught very little. Catches in the small mesh nets differed by including a number of small-bodied species, Notacanthus chemnitzi, Nezumia bairdi, Gaidropsarus ensis and Sebastes sp., while failing to catch any of the larger-bodied R. hippoglossoides.
Catches by longline
The single longline set at 950 m south of Emerald Bank caught primarily C. fabricii and E. princeps (Table 6), as did gillnets at this location. The smallest hooks (10/0) were the most successful in catching these species whereas the largest (14/0) hooks caught only C. fabricii, and in lesser numbers than the smaller hooks despite twice as many 14/0 hooks being set compared to the smaller sizes.
Catches by crab and shrimp traps
Crab and shrimp traps were fished at four or five depths in both locations, although in small numbers (Table 7). Trap catches were composed in large part of species not taken by gillnets and longlines. Coryphaenoides armatus and Anarhichas denticulatus were caught only in crab traps (except for one C. armatus in a shrimp trap). Shrimp traps provided the only records of Myxine glutinosa and accounted for the great majority of Simenchelys parasitica catches (61 of 68 fish, five more coming from crab traps and one each from gillnets and longlines).
Discussion
The primary objective of the present exploratory deep-water fishing operation was to test-fish a wide variety of fixed gear types and sizes, but the main gillnet gear tested (15 cm mesh), and crab and shrimp traps, were fished in general conformity with an area/depth stratified sampling design. This allows broad inferences to be made about the composition and spatial distribution of the demersal fish fauna on the slope off Nova Scotia that is available to fixed gears. The other fixed gear fishing experiments that have been conducted in deep water in the Northwest Atlantic were in more northern areas, Davis Strait (Jørgensen, 1995) and Flemish Pass (Murua and de Cárdenas, 2005), and these used longlines only (a gear not deployed successfully during the present trials).
Variations in the catches of 15 cm gillnets were observed between areas and among depths sampled. There were differences between areas in the relative abundance of species, in species catch rates and in the average sizes of species. These areas are separated by only 4° of longitude and the amount of sampling was minimal, so the differences noted could be largely a reflection of sampling variation. Nonetheless, the areas are topographically different, the eastern sampling area being at the mouth of The Gully, the largest canyon on the east coast of North America (designated by the Canadian Government as a protected area in 2004), whereas south of Emerald Bank sampling was in an area of open slope (and the few near bottom temperature observations made suggest that the water was slightly warmer at fishing depths in the latter area). It is possible therefore that present results do indeed reflect local differences in faunal characteristics. With depth, total numbers caught declined but total weight caught increased. This reflected changes in species composition, the small-bodied species that occurred in abundance at the shallowest stations being replaced at deeper stations by less abundant but larger-bodied species.
Gillnets of sizes other than 15 cm were not fished frequently enough to allow for more than the following speculations about how catches in these might differ. Indications were that gillnets smaller than 15 cm would catch a greater diversity of small-bodied species, whereas nets with meshes larger than 15–18 cm would catch progressively fewer of the same species caught in these nets.
The crab and shrimp traps were fished to determine the abundance of various crustacean species that had been identified by Pohle et al. (1992) as potential candidates for commercial exploitation, but almost no invertebrate species were caught in these other than the deep-sea red crab (Chaceon quinquedens) (Halliday and Cooper, 1991), for which there was already an established fishery. However, the data on fish bycatches in these traps provide additional information on the abundance of some species. The species caught in traps were benthic scavengers. While these included specimens of several species that were caught frequently also in 15 cm gillnets, e.g. A. rostrata, traps (shrimp traps in particular) accounted for virtually all catches of the slender-bodied species M. glutinosa and S. parasitica. Although C. armatus also was caught only in traps, all catches were from those set at 2800 m. This species has a minimum depth limit of about 2000 m (King et al., 2006). Thus, its absence from gillnet catches could be due to the failure to fish gillnets successfully below about 1800 m. Anarhichas denticulatus was the only large-bodied species (four fish, 67–104 cm) caught only in traps when traps and gillnets were fished on the same gear string.
All of the species caught during the fixed gear fishing trials described here have been caught previously in otter trawl surveys on the Scotian Shelf slope (Markle et al., 1988; Halliday et al., 2012). However, there were substantial differences between fixed and mobile gears in the relative importance of species in catches, and in species size compositions.
The relative contributions, by weight, of the top five species caught in gillnets are compared in Fig. 3 to those of the top five species in the otter trawl catches made by Halliday et al. (2012). The percentages contributed to the total catch by C. fabricii and R. hippoglossoides were similar between gears. However, H. affinis and C. coelolepis, dominant species in gillnet catches, were of much lesser relative importance in trawl catches. Conversely, Coryphaenoides rupestris, a co-dominant with C. fabricii in trawl catches, and Alepocephalus agassizii, were completely absent from gillnet catches (except that the one unidentified alepocephalid could have been an A. agassizii).
|
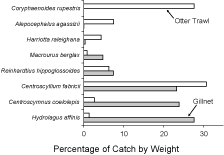 Fig. 3. The relative contributions, by weight, of the top five species caught in all gillnets compared to the relative contributions of the top five species to otter trawl catches made on the Scotian Shelf Slope at similar depths (Halliday et al., 2012) Fig. 3. The relative contributions, by weight, of the top five species caught in all gillnets compared to the relative contributions of the top five species to otter trawl catches made on the Scotian Shelf Slope at similar depths (Halliday et al., 2012)
|
Mean, minimum and maximum lengths of the six species caught in most abundance in 15 cm gillnets are compared with the same statistics for catches of these species in otter trawls (Table 8). Mean lengths were greater in gillnet catches than in small mesh otter trawl catches in four cases. Minimum sizes of these four species in gillnet catches were substantially higher than in otter trawl catches, as would be expected, but maximum sizes were also higher. However, mean sizes of M. berglax and R. hippoglossoides were similar between gears, likely indicating a scarcity of small fish of these species on the Scotian Shelf slope (Halliday et al. 2012).
Despite these differences in species importance, and in species size compositions, between gillnet and otter trawl surveys, both demonstrate the importance of Chimaeriformes and sharks in the Scotian Slope fauna. These groups comprised about one third of the numbers and almost half the weight in the otter trawl catches of Halliday et al. (2012), and 70% by number and 80% by weight in present gillnet catches. The large mesh size of most gillnets fished in the present study likely causes the relative importance in the fauna of these large-bodied species to be over-emphasized. Conversely, the paucity of records of these species in otter trawl surveys of the slope fauna in adjacent areas (Markle and Musick, 1974; Haedrich et al., 1975; Sulak, 1982; Snelgrove and Haedrich, 1985) is likely attributable to a low vulnerability to the small trawls used.
Acknowledgements
Chris Cooper (of the Industry Services and Aboriginal Fisheries Branch, DFO Maritimes Region when the work was conducted) facilitated this project and Gerhard Pohle, Atlantic Reference Centre, Huntsman Marine Science Centre, St. Andrews, N.B., provided information on invertebrate species catches. We are grateful also to D. Kulka, Northwest Atlantic Fisheries Centre, St. John's, Newfoundland and L. Van Guelpen, Atlantic Reference Centre, Huntsman Marine Science Centre, St. Andrews, New Brunswick, for comments on an earlier draft.
References
Bérubé, M., H. Bourdages and A. Fréchet. 2000. Effects of soak time on catch per unit effort using longline and gillnets for the Northern Gulf of St. Lawrence cod stock. Canadian Science Advisory Secretariat (CSAS) Research Document, 2000/150: 28 p.
Haedrich, R. L., G. T. Rowe and P. T. Polloni. 1975. Zonation and faunal composition of epibenthic populations on the continental slope south of New England. J. Mar. Res., 33: 191–212.
Halliday, R. G. and C. Cooper. 1991. Exploration of deepwater resources off the Scotian Shelf. DFO Scotia-Fundy Region, Halifax, N.S., Project Summary, No. 32, Dec. 1991: 4 p.
Halliday, R.G., L. Van Guelpen and D. E. Themelis. 2012. Demersal fish fauna of the continental slope off Nova Scotia, Canada, based on exploratory bottom trawl surveys in 1994–95. J. Northw. Atl. Fish. Sci., 44, 41–60.
JØrgensen, O. A. 1995. A comparison of deep water trawl and long-line research fishing in the Davis Strait. In: Deep-water fisheries of the North Atlantic oceanic slope, A. G. Hopper (ed.) Kluwer Academic Publishers, p. 235–250.
King, N. J., P. M. Bagley and I. G. Priede. 2006. Depth zonation and latitudinal distribution of deep-sea scavenging demersal fishes of the Mid-Atlantic Ridge, 42° to 53°N. Mar Ecol Prog Ser., 319: 263–274. http://dx.doi.org/10.3354/meps319263
Markle, D. F., M. J. Dadswell and R. G. Halliday. 1988. Demersal fish and decapod crustacean fauna of the upper continental slope off Nova Scotia from LaHave to St. Pierre Banks. Can. J. Zool., 66: 1952–1960. http://dx.doi.org/10.1139/z88-286
Markle, D. F. and J. A. Musick. 1974. Benthic-slope fishes found at 900 m depth along a transect in the western N. Atlantic Ocean. Mar. Biol., 26: 225–233. http://dx.doi.org/10.1007/BF00389253
Murua, H. and E. de Cárdenas. 2005. Depth-distribution of deepwater species in Flemish Pass. J. Northw. Atl. Fish. Sci., 37: 1–12. http://dx.doi.org/10.2960/J.v37.m563
Pohle, G., T. J. Kenchington and R. G. Halliday. 1992. Potentially exploitable deepwater resources off Atlantic Canada. Can. Tech. Rep. Fish. Aquat. Sci., 1843: 85 p.
Snelgrove, P. V. R. and R. L. Haedrich. 1985. Structure of the deep demersal fish fauna off Newfoundland. Mar Ecol. Prog. Ser., 27: 99–107. http://dx.doi.org/10.3354/meps027099
Sulak, K. J. 1982. A comparative taxonomic and ecological analysis of temperate and tropical demersal deep-sea fish faunas in the western North Atlantic. Ph.D. dissertation, Rosenstiel School of Marine and Atmospheric Science, University of Miami, Miami, Florida, USA, 211 p.
|