J. Northw. Atl. Fish. Sci., Vol. 56: 11–29
Elizabeth M.T. Bateman1 , Danni L. Harper2 , Tyler Byrne3 , Adam Comeau3 ,
Cornelia E. den Heyer4 , Michael J.W. Stokesbury1
1Department of Biology, Acadia University, Wolfville, NS, Canada
(corresponding author; ElizabethBateman@acadiau.ca), (Michael.Stokesbury@acadiau.ca)
2Population Ecology Science Division, Fisheries and Oceans Canada, Dartmouth, NS, Canada
(Danni.Harper@dfo-mpo.gc.ca),
3Coastal Environmental Observation Technology and Research,
Dalhousie University, Halifax, NS, Canada
(TylerByrne@dal.ca), (Adam.Comeau@dal.ca)
4Ocean Ecosystem Science Division, Fisheries and Oceans Canada, Dartmouth, NS, Canada
(Cornelia.denHeyer@dfo-mpo.gc.ca)
Bateman, E.M.T., Harper, D.L. , Byrne, T., Comeau, A., den Heyer, C.E., and Stokesbury, M.J.W. 2025. Movement and behaviour of Atlantic Halibut (Hippoglossus hippoglossus) in a juvenile hotspot detected by a passive moored acoustic receiver grid and active wave gliders. J. Northw. Atl. Fish. Sci., 56: 11–29. https://doi.org/10.2960/J.v56.m753
Abstract
Novel acoustic receiver applications have recently enabled the monitoring of movements of migratory oceanic species. One such species is the Atlantic Halibut, a commercially important deep-water flatfish that exhibits complex migratory behaviours throughout the Northwest Atlantic Ocean. Here we set up a 144 km2 receiver grid (n = 24 VR4 receivers) in an important halibut hotspot on the Scotian Shelf and tagged 245 halibut with V13 and V16 tags from 2020–22. Receiver performance was assessed using a Receiver Efficiency Index, indicating that the deeper eastern side of the array was an area of high relative importance for future receiver deployments. Many halibut remained in consistent, localized areas over several years, occasionally making short movements into deeper water. We were also able to assess the post-release behaviour of four recaptured pre-tagged halibut within the grid array, indicating a brief period of hyperactivity and an eventual return to their previously observed behaviours. Wave gliders surveyed the grid site annually from 2021–23 to compare the efficacy of active tracking to stationary arrays for future halibut telemetry projects. Two of three tracking missions were disrupted by environmental disturbances, but results from the 2023 tracking mission indicated that the glider was able to detect more individual halibut than the stationary array in the same time frame. However, the short duration of the glider missions precluded their ability to identify movement and migratory patterns. Here we report lessons learned to streamline future project design for open ocean telemetry and halibut acoustic tracking.
Keywords: Acoustic telemetry, Scotian Shelf, commercial fishery, fish tracking
PDF
Download Citation Data

Citation to clipboard
 Reference management software (Endnote, Mendeley, RefWords, Zotero & most other reference management software)
Reference management software (Endnote, Mendeley, RefWords, Zotero & most other reference management software)
LaTex, BibDesk & other specific software
Introduction
Acoustic telemetry has revolutionized the way researchers collect movement data on aquatic species and has ushered in a “golden era” of animal tracking (Hussey et al., 2015; Hays et al., 2019). This technology has since grown as a fisheries conservation tool and provides data that are used to address a growing diversity of management objectives (Matley et al., 2022), including informing fishery quota decisions (Brownscombe et al., 2019), delineating protected areas (Hussey et al., 2017; Martín et al., 2020), and determining rates of natural mortality (Whitlock et al., 2022).
Receiver array geometry is a key consideration for studies addressing management issues because acoustic receivers define the observable study area for tagged organisms. As a result, studies typically monitor aquatic animals in geographically confined systems such as lakes (Hayden et al., 2014; Futia et al., 2024), rivers (Andrews et al., 2018; Burns et al., 2023), and estuaries (Stokesbury et al., 2016; Hollema et al., 2017) to maintain receiver proximity. Similarly, monitoring animals that traverse predictable marine routes through bottlenecks such as Atlantic Bluefin Tuna (Thunnus thynnus) migrations through the Cabot Strait (Block et al., 2019) is also a common approach to maximize detectability.
Receiver arrays are often arranged into a series of lines or “gates” that bisect corridors (Lacroix et al., 2004) and, if paired, can provide a measure of directional movement (Jackson, 2011). Grid deployments, while less common, can help determine fine scale habitat use in a consistent and unbiased way while allowing researchers to distinguish between unused habitat and the absence of data (Krauset al., 2018). Grid arrays are often positioned in land-bound areas like bays to best monitor patterns of residency and departures of tagged individuals (Hussey et al., 2017; Kraus et al., 2018). Grid deployments are occasionally used in open ocean regions to monitor areas of known ecological significance for the study species, such as known spawning locations (Zemeckis et al., 2014). However, there is no configuration of acoustic receivers that is universally appropriate for all studies or locations.
Acoustic telemetry can be logistically challenging for tracking highly mobile species in open ocean settings, particularly when their migratory behaviours and pathways are poorly understood (Jacoby and Piper, 2025). Increasingly, drifters (Sanderson et al., 2023) and mobile autonomous vehicles such as wave gliders are being fitted with acoustic receivers and used to actively track tagged animals (Lembke et al., 2018; Ennasr et al.,2020; Masmitja et al., 2020; Cypher et al., 2023), providing flexible monitoring coverage beyond the scope of traditional fixed arrays. These new technologies are beginning to provide critical information on migratory oceanic fishes such as Atlantic Halibut (Hippoglossus hippoglossus L., 1758).
The Atlantic Halibut is a large and long-lived flatfish found in the boreal and subarctic waters of the Atlantic Ocean (Trumble, 1993). Atlantic Halibut were historically fished to near collapse but have since recovered to healthy abundances in Canadian waters (COSEWIC, 2011; Trzcinski and Bowen, 2016). This is currently Atlantic Canada’s most valuable groundfish species (Fisheries and Oceans Canada, 2025), and considerable efforts have been made by regulatory bodies and industry partners to increase ecological research and establish proactive management strategies. Extensive mark-recapture and satellite tagging of Atlantic Halibut in the Northwest Atlantic (NWA) has revealed that they exhibit diverse migratory behaviours (or “contingents”; Secor, 1999; Ransier et al., 2024) and generally spawn in the winter in deep, continental shelf slope regions (den Heyer et al., 2012; Armsworthy et al., 2014; Kersula and Seitz, 2019; James et al., 2020; Ransier et al., 2024).
Several key knowledge gaps surrounding Atlantic Halibut movement remain, because while informative, these prior methods of tagging have limitations. Conventional tagging only reveals the net movement of a halibut between two points and is limited to areas and times where the fishery operates. Satellite tagging has so far occurred only on adult halibut large enough to support the large, cumbersome tags, therefore only representing the commercially exploitable size class (> 81 cm). Furthermore, satellite tags are restricted to short deployment lengths, so they are not well suited to monitor multi-year site fidelity. This project was established to address these knowledge gaps and was the first to track Atlantic Halibut (hereafter “halibut”) using acoustic telemetry in Canada. Acoustic tags are much smaller than satellite tags with long battery life spanning up to a decade. Tracking challenges included setting up acoustic monitoring infrastructure in deep, borderless habitats and contending with the diverse and wide-ranging migratory behaviours of halibut that are not well established. To deal with these challenges, a large, non-overlapping offshore receiver grid was deployed in an important juvenile halibut hotspot to provide a consistent measure of presence and residency over a large area. The extent of our monitoring capability was expanded through a collaboration with the Ocean Tracking Network (OTN; O’Dor and Stokesbury, 2009) which allowed us to detect halibut migration and dispersal beyond our study site. We paired this stationary grid with autonomous wave glider tracking to assess the efficacy of active tracking for future halibut telemetry projects in regions with or without moored receivers. In this paper, we investigate 1) the efficiency of our open-ocean grid array, 2) the movement and behaviours of tagged halibut within the array, and 3) a comparative assessment using mobile glider-based tracking for monitoring halibut.
Methods
Study site
This study took place in the Gully region on the Scotian Shelf off Nova Scotia, Canada (Fig. 1). The Gully is the largest submarine canyon on the east coast of North America measuring >40 km long, 16 km wide, and with depths >2 500 m (Rutherford and Breeze 2002), part of which became Atlantic Canada’s first Marine Protected Area in 2004. The Gully region has highly complex physical oceanographic processes due to the bathymetrical features of the canyon, highly variable tides, and influences from both the continental shelf and slope hydrodynamics (Han et al. 2002). North of the Gully lies a shallower basin known as the Trough, located between Banquereau and Sable Island Bank approximately 40 km east of Sable Island, Nova Scotia (Lat 43.980945°, Long -59.033243°; Northwest Atlantic Fisheries Organization [NAFO] Division 4Vs; Fig. 1). This area funnels down into the Gully and is a site of persistently high juvenile halibut abundance (Boudreau et al., 2017). The northeastern tip of the Trough was the deployment site for the Halibut Bio-Tracking (HaliBT) array (Fig. 1). Approximately 10 km to the south of this location lies a line of receivers ~17 km across that bisects the northern border of the Gully Marine Protected Area (GULMPA), and a final cluster of receivers that is part of the OTN Sponge Ground Landers array (SPONGE; Fig. 1) is located a further ~27 km down into the protected area of the canyon.

Fig. 1
Acoustic receiver deployment
In September 2020, 25 VR4-UWM acoustic receivers (Innovasea Systems, Inc) were deployed from a chartered vessel in the northeastern tip of the Gully at the Trough (Lat 44.35398, Long -59.12491), 65 km northeast of Sable Island (Fig. 1). The receivers were deployed to construct a 5 by 5 square grid with units spaced at equidistant 3 km intervals creating a 12 km by 12 km monitoring array covering ~144 km2. Due to logistical challenges at sea, one receiver was not deployed, leaving a gap in receiver coverage in the northeastern corner of the grid (Fig. 1). Receivers were each moored by a 91 kg steel plate anchor attached to an Innovasea Ascent™ Acoustic Release 2 (AR2) with 5/16″ Dyneema rope. Moored receivers floated 2 m above the seafloor suspended by an 18″ diameter rigid trawl float providing 26 kg of buoyancy. When triggered, the AR2s allowed the receivers to float to the surface at the end of the study, leaving the sacrificial mooring behind. The array spanned depths from 86–210 m and was retrieved on 31 May 2024 after ~3 years and 8 months of deployment.
Sentinel and range test tag deployment
Sentinel V13-1H tags (n = 5; 60–180 s ping rate, 152 db) were deployed in June 2021 within the array using moorings that suspended them 6 m above the seafloor. These tags provided continuous reference points for detection range for both fixed receivers and mobile glider-mounted receivers. In June 2022, temperature-sensing V13T-1H tags (n = 6, 60–180 s ping rate, 152 db) were deployed 3 m above the seafloor throughout the array at ~20 m depth increments (86–208 m) near receivers to transmit temperature measurements at depth to produce a temperature profile of the seafloor across the bathymetric grade of the area.
Additionally, a range test was set up to the west of the HaliBT array using Innovasea’s V16-6x tags configured with a fixed transmission delay. In 2021, two range test stations were deployed from a research vessel at 221 m and 95 m depth. These were later approached by the glider in a star-shaped pattern. In 2022, two new range test tags were deployed from a fishing vessel in 210 m and 135 m depth for the glider to conduct range tests in a spiral pattern.
Bathymetry and mapping
Ten metre resolution multibeam bathymetry data for the HaliBT array were accessed from the Canadian Hydrographic Service Non-Navigational (CHS NONNA) bathymetric data portal. These data were interpolated to produce contours in ReefMaster (ReefMaster Software Ltd., 2021) then rasterized and plotted in ArcGIS Pro (ESRI 2024). Maps were made in ArcGIS Pro (ESRI, 2024) and R (R Core Team, 2024), the latter using bathymetric data from the marmap package (Pante and Simon-Bouhet, 2013).
Atlantic Halibut capture and tagging
Atlantic Halibut were captured from a chartered commercial fishing vessel employing commercial bottom-longline gear in the HaliBT array. Halibut between 40–65 cm FL (fork length) were surgically tagged with V13-1H acoustic tags while halibut > 65 cm FL were tagged with a larger V16-4H tag (See Table 1 for tag specifications). All halibut were also tagged with two Floy tags as an external marker. Tagged halibut were then released immediately near their capture location. Tagging occurred in September of 2020 and June of 2021 and 2022.

Table 1
Telemetry data acquisition
Data from moored acoustic receivers were retrieved through a remote download link using a benthos VM4 modem towed behind a Liquid Robotics SV2 or SV3 autonomous wave glider sub (Fig. 2) operated by Coastal Environmental Observation Technology and Research (CEOTR). Data passed to the glider from the benthos modem were sent back to shore over the Iridium Router-Based Unrestricted Digital Internetworking Connectivity Solutions (RUDICS) satellite network and were also stored internally on the glider. Glider download missions occurred annually in the autumn from 2021–23.

Fig. 2
Acoustic telemetry data were also collected via mobile glider tracking using a Liquid Robotics SV3 wave glider (Fig. 2). In 2021, the glider was equipped with two Vemco Mobile Transceivers (VMT; one mounted on the bottom of the float, the other on the bottom of the sub) and one VR2C (mounted on the sub) for the tracking surveys. In 2022, the glider was equipped with 1) the prototype Innovasea Mobile-RX receiver with the head oriented just forward of the bow of the sub, 2) a VMT, and 3) a VR2C positioned at the aft end of the sub with the head pointed downward. In 2023, the glider was equipped with two VMTs on the sub (one pointing up and the other pointing down) and a VM4 mounted on a towfish, pointing downward (Fig. 2). The towfish was towed between 10–12 m behind and 7–10 m below the sub. The glider recorded detections of tagged halibut in the study area along pre-programmed North-South and East-West “lawnmower” pattern transects travelled annually from 2021–23.
HaliBT array receiver efficiency
In ideal conditions in deep water with low turbidity, sandy substrate, and low currents, transmission signals from the highest-powered tags available (V16-5H) can propagate to 1 200 m (Loher et al., 2017). We did not pursue a dedicated receiver range test in our study area due to the challenging logistics of this location, however, the transmission range in our study was suspected to be lower due to our lower-powered tags, the complex bathymetry in the HaliBT array, and the potentially strong tidal currents occurring in the Gully. Another method of assessing the performance of an array is the Receiver Efficiency Index (REI), introduced by Ellis et al. (2019). This index is a composite measure of relative importance of each receiver in an array, incorporating ratios of detections and individuals at each receiver, while also inherently incorporating differences in receiver range (Kendall et al., 2021). The resulting indices can help identify redundant or low-value stations to inform future array configurations that will maximize efficiency while allowing an array to be maintained with fewer stations (Ellis et al., 2019). We used the following equation developed by Ellis et al. (2019):
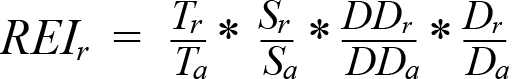
where r is an individual receiver and a is the whole array, T = number of tag IDs in the study, S = number of species, DD = detection days, and D = number of days a specific receiver was deployed within the array. Given that halibut are the only species of interest for the study and that all receivers have the same deployment period, both Sr/Sa (the proportion of species in the array detected at each receiver) and Dr/Da were set to 1.
Detection data analysis and visualization
Detection data downloaded from the HaliBT array and collected by the wave gliders were accessed through the OTN data portal. Detection data and project metadata were managed and visualized in the R programming environment RStudio (R Core Team, 2024).
Glider tracking analysis
Glider tracks and glider-based detections of tagged halibut were superimposed in R. The timing of each glider transect pass (e.g., North-South, East-West) and the associated detections were limited to when the glider was in the array conducting the dedicated tracking surveys. To compare relative detection rates between the mobile glider and the stationary receiver array, detections made by the HaliBT array were subset to only include those which overlapped temporally with glider occurrence in the fixed array. Number of detections and unique tag IDs detected were tallied for both tracking methods for the duration of each directional survey.
Glider range test
The maximum distance at which the glider detected each sentinel tag deployed within the fixed array was calculated in ArcGIS Pro. Since sentinel tags were the same model and configuration as the smallest halibut tags, this was considered a within-array glider range test proxy and an appropriate measure of detection distance of the halibut.
Results
Atlantic Halibut length distribution
Fork lengths of tagged halibut ranged from 50 cm to 141 cm and the mean varied by tagging year (Table 2).

Table 2
Detections by yearly release group
Seven halibut tagged in September 2020 were captured and released on Banquereau, outside the study area. Five of those halibut were not detected on the array. Of the two individuals detected, one large halibut (110 cm) passed through the HaliBT array briefly on 23 Dec 2020, then again on 24 Jan 2021. The second smaller halibut (69 cm) was not detected until over two years after release when it was observed in the array over two days in January 2022 (Table 3; Fig. 3).

Table 3
The 2021 release group (n = 138) was at large for approximately 2 years and 11 months as of the May 2024 HaliBT array retrieval mission (Table 1). Transmissions of V13 tags (n = 70) were projected to have ceased by March 2023, but likely stopped transmitting in April 2023 based on available data. V16 tags (n = 68) are projected to continue transmitting until February 2028 (Table 1). Four halibut from this release group have been reported harvested, one by the DFO groundfish survey trawl in July 2023 near the HaliBT array, one at an unknown time and location in early 2023 by a fishing vessel, the third on the continental shelf edge below Sable Island in March 2024, and the fourth off the coast of Saint Pierre and Miquelon in May 2024.

Fig. 3
The 2022 release group (n = 100) was at large for approximately 1 year and 11 months as of the May 2024 HaliBT array retrieval mission. Transmissions of V13 tags (n = 50) ceased in March 2024 and transmissions of V16 tags (n = 50) are projected to stop by February 2029. Two tagged halibut from this group were reported to have been harvested at an unknown time and location in early 2023 by fishing vessels.
All but five halibut in the 2021/2022 release groups were detected by the HaliBT array at least once following release (Table 3). Numbers of tagged halibut present in the array varied month by month with peaks in the summer (exclusive of tagging efforts) and lows in the winter (Fig. 3). Halibut display a variety of migratory behaviours which may account for the variability in the number of detections and the number of days each individual halibut was detected in the array. A subgroup of halibut was present within the HaliBT array year-round, termed residents.
Glider tracking
Glider tracking missions had variable success over the years. In 2021, the glider had to be manoeuvred off course multiple times to avoid vessels and approaching hurricanes. Upon the glider’s return to the HaliBT array to begin the final transect, navigational control was lost and the glider drifted into the Gully MPA where it was retrieved. Extensive damage to the thruster and rudder complex resulting from a shark bite rendered the unit disabled.
In 2022, the first North-South glider transect was interrupted when it was re-routed to avoid the approach of the Category 3 Hurricane Fiona. Hurricane Fiona transitioned to a large, powerful post-tropical storm upon arrival in Atlantic Canada with record-breaking drops in barometric pressure. In the turbulence on 24 September 2022, the glider’s umbilical tether detached, losing both the sub and all receiver packages. Sensor readings are unreliable following peak hurricane activity, but at the time of the incapacitation, the glider measured ~ 15 m wave heights and 111–148 km/h winds. The catastrophically damaged glider float drifted north and was retrieved, but no acoustic detection data were salvaged from this mission.

Fig. 4
In 2023, the glider had to be routed out of the array during the first East-West transect to avoid the approaching Hurricane Idalia. When wave heights exceeded 4 m during the first North-South transect, the glider was held in a bowtie course to best handle the energetic sea state (Fig. 4). Other deviations from the transects during the survey occurred when the glider was avoiding vessel traffic, but otherwise, the survey was completed without major issues. The survey ran from 28 August 2023 to 5 October 2023 and consisted of a total of 42 East-West passes and 42 North-South passes, averaging 1.0 knot with a maximum of 3.1 knots attained during that time. The downward-pointing VM4 logged most of the detections (n = 1 624) and the downward-pointing VMT logged only 372 detections during the tracking transects (Fig. 4). The upward-pointing VMT logged no detections, likely due to its proximity to and orientation toward surface turbulence.
The glider, performing a “lawnmower” search pattern, consistently detected more individual halibut tag IDs than the HaliBT array during each transect period, but the HaliBT array made more detections overall during the same period, likely due to the simultaneous recording of its 24 receivers and consistent detection of nearby halibut that may have undertaken little movement for some time (Table 4). However, the HaliBT array occasionally detected halibut that were not detected by the glider.

Table 4
Eighteen halibut were detected by the glider in all four survey transects, but only eleven of those halibut were also detected by the HaliBT array in that time (Fig. 5). This disparity indicates that the gaps between receivers in the HaliBT array allow many halibut to occupy space in the array where they are undetected, but also that these gaps can be filled in on occasion by glider tracking.

Fig. 5
HaliBT array receiver efficiency
REI values were highest in the eastern side of the array, with station 15 (Lat 44.35413, Long -59.04956; 142 m depth; Fig. 6) having the highest index of 0.583 (Table 5). The high indices in this region likely relate to denser and more resident aggregations of halibut. Stations with REI > 0.2 may be the best candidates for maintaining receivers in future tracking efforts. Most glider detections of halibut occurred in similar locations as the HaliBT receivers with the highest REI values (Fig. 4; Fig. 6). This may indicate that the eastern side of the array supports a local abundance of halibut and not just high REI values resulting from better acoustic transmission conditions than other areas. Although neighbouring receiver stations were only 3 km apart, the REIs of stations immediately surrounding station 15 dropped by half or more, possibly indicating particularly high detection range at station 15. Halibut were detected on the most days at station 15 (604 total detection days, 95% of entire monitoring period), whereas they were detected on the fewest days at station 23 (Lat 44.40825, Long -59.12527; Fig. 6) at 105 m (18 total detection days, 3% of monitoring period, Table 5).

Fig. 6
Station 6 (Lat 44.32701, Long -59.049548; 166 m depth; Fig. 6) was found washed up on Sable Island (~65 km southwest of the original receiver location) on 11 April 2023. The last detection logged by the unit occurred on March 27, 2023. REI was only calculated until the end of March 2023 to best represent when the entire array was still in place.
Glider detection range
The SV3 wave glider’s detection range was highly variable. The distances at first detection of the sentinel tags within the array varied by location, the highest being 893 m at the deepest station (208 m) and the lowest being 553 m at the shallowest station (86 m; Fig. 7).

Table 5
Halibut movement and habitat use in the HaliBT array
Halibut were generally only detected on a small number of HaliBT receivers out of the available 24 (Table 3), indicating discrete use of the study area. The regions of the array that were most visited by halibut were around the shallow point extending down the middle rather than the deepest station, suggesting that the halibut aggregated around major structure throughout the study period (Fig. 6). Many halibut were transient and/or seasonal visitors to the HaliBT array, but some displayed resident behaviour, enabling a closer look into spatial use of the area. Residents remained in the Gully region year-round, with most generally staying in the vicinity of the HaliBT array. The locations of 18 residents from the 2021 release group were plotted as an example due to the long tag deployment time and their tendency to stay within the array (Fig. 8). Halibut in the 2021 resident group tended to remain in one consistent location for extended periods but often made short forays south and then back (Fig. 8). Very few halibut ventured into the shallowest northern edge of the array (receiver stations 21–24; Fig. 8). Even halibut that were not necessarily resident to the area remained relatively stationary when present, as evidenced by glider tracking (Fig. 5). Eighteen of the halibut were detected during every transect during the 2023 glider survey, all of which showed little to no movement during the 39-day survey period (Fig. 5). All of the eighteen halibut were active over the years, including making seasonal movements and venturing deeper into the Gully as monitored by the GULMPA receiver line (Fig. 9).

Fig. 7
The HaliBT array provided an unexpected opportunity to assess catch-and-release (C&R) survival and post-release movement. On 22 June 2022, during the final tagging trip of the project in the HaliBT array, four halibut were caught on the longline and identified as previously tagged individuals based on their healed incisions and Floy tags. Those halibut ranged from 53–82 cm at the time of tagging and all followed the expected growth rate of ~10 cm/year (Fisheries and Oceans Canada, 2020), suggesting they were in good health. These halibut were handled the same way as they are in the commercial fishery (e.g. gaffed through the mouth, brought on deck, and measured) and once their Floy tag information was recorded, they were released overboard as would halibut under the commercial retention size (~ 5–8 minutes total handling time).
In the month leading up to their capture, all four halibut were detected only at station 15. After C&R, tag ID 47676 (seasonal migrant showing summer site fidelity to the HaliBT array; 76 cm at time of tagging) was not detected anywhere for 19 days until it was detected by an external receiver line at the border of the GULMPA 17 km away. This halibut then returned to the HaliBT array to resume activity between several stations (Fig. 10 B). Similarly, tag ID 47 644 (resident, 53 cm at time of tagging) was undetected post-release for 15 days, then was detected regularly back at station 15. Tag ID 2716 (seasonal migrant to the HaliBT array; 82 cm at time of tagging) was detected in consistent 6-day intervals in the month leading up to the C&R event, then went undetected for 25 days until it was detected again on several stations in the HaliBT array. Tag ID 47 635 (resident, 60 cm at time of tagging) was detected at station 6 immediately south of station 15 after release, then went undetected for 13 days until it was detected on one day by the GULMPA line. It was detected again by the GULMPA line nearly three months later, then back in the HaliBT array resuming regular resident behaviour. A similar gap in detection occurred in 2021 after this individual was first captured, tagged, and released (Fig. 10 A).

Fig. 8
Discussion
In this study, we examined the efficacy of using an open ocean receiver grid and paired wave gliders to track halibut on the Scotian Shelf while comparing both methods in situ. The grid receiver array positioned over a known halibut aggregation area proved to be a successful method of monitoring multi-year halibut behaviour. The large spatial coverage of the array revealed that the deeper eastern channel was an area of higher halibut density, as described by the commercial fishermen who suggested that the tagging efforts be carried out in that location. These observations may help guide future localized studies of habitat ecological linkages to substrate type and prey composition. This array design facilitated a unique opportunity to assess the effectiveness of mobile glider tracking, observe small-scale movements of halibut for extended periods of time, and observe how halibut behave post catch-and-release.

Fig. 9
Efficacy of an open-ocean receiver grid
The HaliBT array collected >900 000 detections of tagged halibut and provided important and detailed data on the poorly described movement behaviours of halibut, both on the scale of a few kilometers and on a broader scale when connected with other nearby receiver arrays in the Gully ecosystem and other areas through the OTN. Though this type of array design is particularly effective for fish that display high site fidelity like halibut, the HaliBT array also detected tag IDs associated with 14 other acoustic tracking projects linked through the OTN that monitored a variety of migratory pelagic species including Atlantic Bluefin Tuna, Atlantic Salmon (Salmo salar), and Leatherback Sea Turtles (Dermochelys coriacea). This underscores the broader application that large oceanic grid arrays can offer to the collaborative tracking community.
Feasibility of mobile glider tracking
Autonomous gliders travelling in set transects over the HaliBT array detected a higher number of acoustically tagged halibut than did the fixed array in the same time frame. Similar results were found in another offshore groundfish study that used gliders to augment fixed arrays (Zemeckis et al., 2019). However, several halibut that were detected by the fixed array were not detected by the glider.

Fig. 10
The continual tracking over the study area for approximately one month emphasized how localized halibut can be for extended periods and provided finer-scale detections than were available from the widely spaced array. Using sentinel tags at different depths in the array, we found that the detection range of the glider varied considerably by up to 340 m between locations. Considering that the largest detection range was at the deepest station and the smallest detection range occurred at the shallowest station, there are likely many factors affecting the glider’s detection range, including wave action and other ambient noise (How and De Lestang, 2012) and thermocline structure (Huveneers et al., 2016). The detection range we report here is specific to the short time that the glider was in the array and would likely change based on seasons and the physical oceanography at a given time. Although we did not explicitly investigate these relationships in this study, environmental conditions that could impact sound transmission such as thermocline displacement (O’Brien and Secor, 2021) and wind-driven turbulence (Gjelland and Hedger, 2013) should be taken into consideration in future projects. In the 2023 glider survey, most detections were made by the tow-behind VM4 unit which was oriented pointing downward. This orientation biases the VM4 to pick up signals from below, which was likely advantageous in our study as halibut are generally benthic, but this configuration may not be as effective for tracking pelagic fishes.
In addition to the lawnmower-pattern surveys used in our study, the use of gliders can greatly extend the monitoring coverage of a study site by travelling the perimeter of a fixed array or filling in gaps between receiver arrays. Unfortunately, the success of glider missions is unpredictable, as demonstrated by the shark attack in 2021 and the damage inflicted by Hurricane Fiona in 2022, resulting in the loss of several receivers and the collected data. Although gliders are autonomous, weather conditions and glider status must be monitored closely by a team of pilots during surveys. Further, discontinuous and infrequent glider surveys over an area of interest are likely to miss important nuances in fish movement, such as seasonal migration. For example, tagged halibut in this study area that exhibited seasonal migratory behaviour regularly spent extended periods in the HaliBT array, but left in the late fall, likely to migrate to their winter spawning grounds. Using data acquired from the short-term glider surveys alone, it was not possible to distinguish residents from migrants. Given these limitations, glider tracking may not be an appropriate standalone substitute for fixed receiver arrays except in locations with heavy fishing pressure or with protected benthic ecosystems where fixed moorings are not possible. The scope of the research question may also influence the utility of glider tracking, where there would be an advantage for detecting short-term events like spawning aggregations.
Mobile tracking has also been used to identify fish mortalities (i.e., sedentary tag IDs) in tagging studies using both VR100 receivers by boat (Chavarie et al., 2022) and by autonomous wave gliders (Zemeckis et al., 2019) under the assumption that in many cases, cessation of horizontal movement during repeated surveys over the area is indicative of death (Klinard and Matley, 2020). Although identifying mortalities was not a goal of this project, it was noted that many tagged halibut did not move at all during the entirety of the 39-day glider survey, which normally would suggest a mortality event. However, the long-term stationary receiver detections of those same halibut revealed extensive activity in the HaliBT array and beyond but over a much longer timescale than the glider survey. The use of gliders for detecting mortalities may therefore be impractical for flatfish species like Atlantic Halibut that can be both highly mobile and can remain in one place for extended periods.
Catch-and-release survival
Assessing the behaviour and survival of fish following a C&R event is rarely possible independent of tagging effects and tagging mortality and is difficult to do without a well-established receiver array. The recapture of four previously-tagged halibut within the HaliBT array provided a glimpse into the survival and behaviour of halibut post-release, facilitated by the extensive coverage of a grid array. Though the sample size is insufficient to provide an estimate of mortality, the significant number of detections from the tagged halibut in this study suggests that survival of halibut is high following capture, tagging, and release. Similarly, previous studies have found that post-release survival of halibut (both Atlantic and Pacific Hippoglossus stenolepis) captured by longline is high following standard fishing practices and quick handling (Neilson et al., 1989; Kaimmer and Trumble, 1998). The recaptured halibut in this study had good body condition, healed incisions, and normal growth rates a year post-tagging, and the detections following recapture in 2022 indicated survival and an eventual return to regular behaviour. All halibut went undetected for some time after recapture and release and two were detected further down the Gully canyon on the GULMPA line before returning to the HaliBT array. This suggests that halibut might undergo a flight response after release as is documented in other species, such as post-release downstream movements of otherwise upstream-swimming Atlantic Salmon (Mäkinen et al., 2000). Other fish species captured by angling have been found to undergo a period of hyperactivity after release followed by a period of low activity (Cooke et al., 2000; Gurshin and Szedlmayer, 2004). Catch-and-release effects from angling have been previously assessed in Atlantic Halibut using electronic tags (Ferter et al., 2016), but the small sample size, high rates of external tag shedding, and the assumption that mortality is defined by a cessation of vertical movement in that exploratory project limited the conclusions that could be drawn from the results.
The recapture events within the HaliBT array presented the opportunity to monitor the movements of pre-tagged halibut post-release after being captured by longline and handled as they would in a commercial fishery. Although C&R may have elicited a flight response, all halibut eventually returned to their original capture location. Halibut have already been found to display remarkable site fidelity (James et al., 2020), but these detections in the receiver grid demonstrated that halibut returned to their exact location even after a stressful capture event. Furthermore, catch-and-release did not alter the overall migratory behaviour of these halibut, as residents continued to occupy the HaliBT array upon their return and seasonal migrants continued to depart the array during the presumed spawning season, as was found in a similar observation reported for pre-tagged Atlantic Cod (Gadus morhua; Ferter et al., 2015). Given that the Trough and Gully receive considerable fishing pressure (Rutherford and Breeze, 2002) and that this region in particular supports undersized halibut (Boudreau et al., 2017), it is possible that halibut are captured many times before they are legally harvested. However, these few instances of a true C&R event suggest that the short-term survival and overall migratory behaviours of halibut may not be significantly impacted by release from a longline when the fish are in good condition and handling time is short.
Future halibut tracking considerations
The high REI values in the deep eastern channel in the HaliBT array indicate a high relative importance of that area for the tagged halibut in our study and suggest that it may be valuable and effective to maintain deployments of a small number of receivers in that region for future long-term halibut monitoring. This observation was supported by the glider tracking survey, which revealed that most halibut were found on that deep eastern side, thereby indicating that the REI values in this study were not just a matter of detection efficiency but also localized density. In the future, it may be beneficial to test mooring receivers higher in the water column and pointing downward to maintain improved line-of-signal with benthic-oriented tagged fish such as halibut (Long et al., 2023) and to reduce shadowing effects of complex bathymetry.
Glider tracking offers some potential advantages for examining halibut spatial ecology that were not explored in the scope of this project. The number, exact locations, and characteristics of halibut spawning sites are still undescribed in the NWA, which presents a major management challenge. While recent PSAT studies have begun to address the spawning site knowledge gap (see Le Bris et al., 2018; Liu et al., 2019; Ransier et al., 2024), very few archived data packages have been retrieved from the Eastern Scotian Shelf. Acoustic tags present a distinct advantage over PSATs in the ease and cost-effectiveness of deploying more tags with a battery life that may extend up to a decade. Additionally, acoustic tags can be deployed on smaller fish than PSATs, which can reveal the migratory behaviours of that understudied demographic. However, halibut often make use of habitat that is deeper than acoustic receivers can be maintained and sufficient coverage along the edge of the continental shelf slopes where halibut are presumed to spawn is not feasible. Autonomous gliders are powerful tools for searching for tagged fish in the open ocean (Oliver et al., 2013) and can be adaptively routed to follow tagged individuals (Cypher et al., 2023). Many halibut in this project were absent from the HaliBT array during the presumed spawning season but were found to return at regular annual intervals. Given this information, it may be possible to send gliders to search continental shelf edges near the HaliBT array for tagged fish in the winter to determine Scotian Shelf halibut spawning sites. Due to the challenges of operating wave gliders in the winter, namely low light levels and frequent rough conditions, other types of gliders such as the subsurface Teledyne Webb Research Slocum glider could be explored for this purpose.
Conclusion
In recent years, methods of monitoring acoustically tagged species have evolved, allowing researchers to approach their studies in new ways. Open ocean receiver grids can help identify movements of understudied marine species in a small area but are in some cases impractical for wide-ranging transient species. Our study demonstrates how beneficial these grid arrays can be for investigating both the small-scale movements of halibut and their long-term migratory patterns. However, the success of our grid array is due to the decades of data from the commercial fishery and government surveys indicating that our study site is an area of persistent halibut aggregation, and we emphasize that the establishment of open-ocean grid arrays in areas without similar long-term aggregations may not be as successful.
Autonomous wave gliders are not only able to remotely offload data from offshore receivers but can also act as mobile tracking platforms. We found that gliders were able to detect more tagged halibut in the array than the stationary receivers in the same time frame but were unable to determine long-term nuances in halibut behaviour. By presenting the successes and challenges of our monitoring methods, we hope to inform future telemetry research, particularly of halibut and other offshore species.
Acknowledgements
This research was supported by the Atlantic Halibut Council and funded through the Atlantic Fisheries Fund. We thank the crew of the F/V Sleepy Devil for the critical role they played in receiver and tag deployments and their onboard hospitality; Brad Hubley for his contributions to the HaliBT array design; Sue l’Orsa, Keith Spafford, and Tristan Norman for their contributions to the glider tracking and range testing; Dr. Fred Whoriskey for his contributions to the project planning; Dr. Brian Sanderson for his advice regarding glider analysis and bathymetric mapping; Dr. Lingbo Li for her support; Dr. Sam Andrews for his revisions and assistance with ArcGIS; Dr. Alan Clark for his helpful perspectives and advice; and the many students, technicians, and observers who helped in the field over the years. All detections recorded by external receiver arrays were made available through the Ocean Tracking Network.
References
Andrews, S. N., Wallace, B., Gautreau, M., Linnansaari, T., and Curry, R. A. 2018. Seasonal movements of striped bass Morone saxatilis in a large tidal and hydropower regulated river. Environmental Biology of Fishes, 101(10), 1549–1558. https://doi.org/10.1007/s10641-018-0799-y
Armsworthy, S. L., Trzcinski, M. K., and Campana, S. E. 2014. Movements, environmental associations, and presumed spawning locations of Atlantic halibut (Hippoglossus hippoglossus) in the northwest Atlantic determined using archival satellite pop-up tags. Marine Biology, 161(3), 645–656. https://doi.org/10.1007/s00227-013-2367-5
Block, B. A., Whitlock, R., Schallert, R. J., Wilson, S., Stokesbury, M. J. W., Castleton, M., and Boustany, A. 2019. Estimating Natural Mortality of Atlantic Bluefin Tuna Using Acoustic Telemetry. Scientific Reports, 9(1), 4918. https://doi.org/10.1038/s41598-019-40065-z
Boudreau, S. A., Shackell, N. L., Carson, S., and den Heyer, C. E. 2017. Connectivity, persistence, and loss of high abundance areas of a recovering marine fish population in the Northwest Atlantic Ocean. Ecology and Evolution, 7(22). https://doi.org/10.1002/ece3.3495
Brownscombe, J. W., Adams, A. J., Young, N., Griffin, L. P., Holder, P. E., Hunt, J., Acosta, A., Morley, D., Boucek, R., Cooke, S. J., and Danylchuk, A. J. 2019. Bridging the knowledge-action gap: A case of research rapidly impacting recreational fisheries policy. Marine Policy, 104, 210–215. https://doi.org/10.1016/j.marpol.2019.02.021
Burns, C. W., Gibeau, P., Robichaud, D., McCulloch, C., Novoa, J., and Lockert, K. 2023. Eulachon migration patterns in the lower Fraser River revealed through acoustic telemetry. Transactions of the American Fisheries Society, 152(5), 632–649. https://doi.org/10.1002/tafs.10428
Chavarie, L., Honkanen, H. M., Newton, M., Lilly, J. M., Greetham, H. R., and Adams, C. E. 2022. The benefits of merging passive and active tracking approaches: New insights into riverine migration by salmonid smolts. Ecosphere, 13(5), e4045. https://doi.org/10.1002/ecs2.4045
COSEWIC. 2011. COSEWIC assessment and status report on the Atlantic Halibut Hippoglossus hippoglossus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 48 pp. (https://publications.gc.ca/collections/collection_2013/ec/CW69-14-643-2012-eng.pdf).
Cooke, S. J., Philipp, D. P., Schreer, J. F., and McKinley, R. S. 2000. Locomotory Impairment of Nesting Male Largemouth Bass Following Catch-and-Release Angling. North American Journal of Fisheries Management, 20(4), 968–977. https://doi.org/10.1577/1548-8675(2000)020<0968:LIONML>2.0.CO;2
Cypher, A. D., Statscewich, H., Campbell, R., Danielson, S. L., Eiler, J., and Bishop, M. A. 2023. Detection efficiency of an autonomous underwater glider carrying an integrated acoustic receiver for acoustically tagged Pacific herring. ICES Journal of Marine Science, 80(2), 329–341. https://doi.org/10.1093/icesjms/fsac241
den Heyer, C. E., Armsworthy, S., Wilson, S., Wilson, G., Bajona, L., Bond, S., and Trzcinski, M. K. 2012. Atlantic Halibut All-Sizes Tagging Program Summary Report for 2006 to 2011 (2992; Canadian Technical Report of Fisheries and Aquatic Sciences, p. 43). Fisheries and Oceans Canada.
Ellis, R. D., Flaherty-Walia, K. E., Collins, A. B., Bickford, J. W., Boucek, R., Walters Burnsed, S. L., and Lowerre-Barbieri, S. K. 2019. Acoustic telemetry array evolution: From species- and project-specific designs to large-scale, multispecies, cooperative networks. Fisheries Research, 209, 186–195. https://doi.org/10.1016/j.fishres.2018.09.015
Ennasr, O., Holbrook, C., Hondorp, D. W., Krueger, C. C., Coleman, D., Solanki, P., Thon, J., and Tan, X. 2020. Characterization of acoustic detection efficiency using a gliding robotic fish as a mobile receiver platform. Animal Biotelemetry, 8(1), 32. https://doi.org/10.1186/s40317-020-00219-7
ESRI. ArcGIS Pro (Version 3.3). 2024. [Computer software].
Ferter, K., Hartmann, K., Kleiven, A. R., Moland, E., and Olsen, E. M. 2015. Catch-and-release of Atlantic cod (Gadus morhua): Post-release behaviour of acoustically pretagged fish in a natural marine environment. Canadian Journal of Fisheries and Aquatic Sciences, 72(2), 252–261. https://doi.org/10.1139/cjfas-2014-0290
Ferter, K., Rikardsen, A. H., Evensen, T. H., Svenning, M.-A., and Tracey, S. R. 2016. Survival of Atlantic halibut (Hippoglossus hippoglossus) following catch-and-release angling. Fisheries Research, 186, 634–641. https://doi.org/10.1016/j.fishres.2016.05.022
Fisheries and Oceans Canada. 2020. Stock Status Update of Atlantic Halibut (Hippoglossus hippoglossus) on the Scotian Shelf and Southern Grand Banks in NAFO Divisions 3NOPs4VWX5Zc. DFO Can. Sci. Advis. Sec. Sci. Resp. 2020/017.
Fisheries and Oceans Canada. 2025, January 30. Seafisheries landed value by province, 2023 [Statistics]. Fisheries and Oceans Canada, Public Affairs, Digital and Creative Services. https://www.dfo-mpo.gc.ca/stats/commercial/land-debarq/sea-maritimes/s2023pv-eng.htm
Futia, M. H., Binder, T. R., Henderson, M., and Marsden, J. E. 2024. Modeling regional occupancy of fishes using acoustic telemetry: A model comparison framework applied to lake trout. Animal Biotelemetry, 12(1), 25. https://doi.org/10.1186/s40317-024-00380-3
Gjelland, K. Ø., and Hedger, R. D. 2013. Environmental influence on transmitter detection probability in biotelemetry: Developing a general model of acoustic transmission. https://besjournals.onlinelibrary.wiley.com/doi/10.1111/2041-210X.12057
Gurshin, C. W. D., and Szedlmayer, S. T. 2004. Short-term survival and movements of Atlantic sharpnose sharks captured by hook-and-line in the north-east Gulf of Mexico. Journal of Fish Biology, 65(4), 973–986. https://doi.org/10.1111/j.0022-1112.2004.00501.x
Han, G., Roussel, P., and Loder, J. 2002. Seasonal-Mean Circulation and Tidal Currents in The Gully. In Coastal and Estuarine Modeling (pp. 22–34). American Society of Civil Engineers. https://doi.org/10.1061/40628(268)2
Hayden, T. A., Holbrook, C. M., Fielder, D. G., Vandergoot, C. S., Bergstedt, R. A., Dettmers, J. M., Krueger, C. C., and Cooke, S. J. 2014. Acoustic Telemetry Reveals Large-Scale Migration Patterns of Walleye in Lake Huron. PLOS ONE, 9(12), e114833. https://doi.org/10.1371/journal.pone.0114833
Hays, G. C., Bailey, H., Bograd, S. J., Bowen, W. D., Campagna, C., Carmichael, R. H., Casale, P., Chiaradia, A., Costa, D. P., Cuevas, E., Bruyn, P. J. N. de, Dias, M. P., Duarte, C. M., Dunn, D. C., Dutton, P. H., Esteban, N., Friedlaender, A., Goetz, K. T., Godley, B. J., … Sequeira, A. M. M. 2019. Translating Marine Animal Tracking Data into Conservation Policy and Management. Trends in Ecology and Evolution, 34(5), 459–473. https://doi.org/10.1016/j.tree.2019.01.009
Hollema, H. M., Kneebone, J., McCormick, S. D., Skomal, G. B., and Danylchuk, A. J. 2017. Movement patterns of striped bass (Morone saxatilis) in a tidal coastal embayment in New England. Fisheries Research, 187, 168–177. https://doi.org/10.1016/j.fishres.2016.11.006
How, J., and De Lestang, S. 2012. Acoustic tracking: Issues affecting design, analysis and interpretation of data from movement studies. Marine and Freshwater Research, 63, 312–324. https://doi.org/10.1071/MF11194
Hussey, N. E., Kessel, S. T., Aarestrup, K., Cooke, S. J., Cowley, P. D., Fisk, A. T., Harcourt, R. G., Holland, K. N., Iverson, S. J., Kocik, J. F., Mills Flemming, J. E., and Whoriskey, F. G. 2015. Aquatic animal telemetry: A panoramic window into the underwater world. Science, 348(6240), 1255642. https://doi.org/10.1126/science.1255642
Hussey, N. E., Hedges, K. J., Barkley, A. N., Treble, M. A., Peklova, I., Webber, D. M., Ferguson, S. H., Yurkowski, D. J., Kessel, S. T., Bedard, J. M., and Fisk, A. T. 2017. Movements of a deep-water fish: Establishing marine fisheries management boundaries in coastal Arctic waters. Ecological Applications, 27(3), 687–704. https://doi.org/10.1002/eap.1485
Huveneers, C., Simpfendorfer, C. A., Kim, S., Semmens, J. M., Hobday, A. J., Pederson, H., Stieglitz, T., Vallee, R., Webber, D., Heupel, M. R., Peddemors, V., and Harcourt, R. G. 2016. The influence of environmental parameters on the performance and detection range of acoustic receivers. Methods in Ecology and Evolution, 7(7), 825–835. https://doi.org/10.1111/2041-210X.12520
Jackson, G. D. 2011. The Development of the Pacific Ocean Shelf Tracking Project within the Decade Long Census of Marine Life. PLOS ONE, 6(4), e18999. https://doi.org/10.1371/journal.pone.0018999
Jacoby, D. M. P., and Piper, A. T. 2025. What acoustic telemetry can and cannot tell us about fish biology. Journal of Fish Biology, 106(5), 1260–1284. https://doi.org/10.1111/jfb.15588
James, T. L., Landsman, S. J., Ramsay, L. L., Giffin, M. D., Le Bris, A., and van den Heuvel, M. R. 2020. Migration patterns of Atlantic halibut captured in the southern Gulf of St. Lawrence as assessed with pop-up satellite archival and Floy tags. Canadian Journal of Fisheries and Aquatic Sciences, 77(7), 1233–1242. https://doi.org/10.1139/cjfas-2019-0262
Kaimmer, S. M., and Trumble, R. J. 1998. Injury, condition, and mortality of Pacific halibut bycatch following careful release by Pacific cod and sablefish longline fisheries. Fisheries Research, 38(2), 131–144. https://doi.org/10.1016/S0165-7836(98)00153-2
Kendall, M. S., Williams, B. L., Ellis, R. D., Flaherty-Walia, K. E., Collins, A. B., and Roberson, K. W. 2021. Measuring and understanding receiver efficiency in your acoustic telemetry array. Fisheries Research, 234. https://doi.org/10.1016/j.fishres.2020.105802
Kersula, M., and A. Seitz. 2019. Diverse migratory behaviors of Atlantic halibut (Hippoglossus hippoglossus, L.) based on the 2000–2017 Maine halibut tagging program. J. Northw. Atl. Fish. Sci., 50: 13–24. https://doi.org/10.2960/J.v50.m719
Klinard, N. V., and Matley, J. K. 2020. Living until proven dead: Addressing mortality in acoustic telemetry research. Reviews in Fish Biology and Fisheries, 30(3), 485–499. https://doi.org/10.1007/s11160-020-09613-z
Kraus, R. T., Holbrook, C. M., Vandergoot, C. S., Stewart, T. R., Faust, M. D., Watkinson, D. A., Charles, C., Pegg, M., Enders, E. C., and Krueger, C. C. 2018. Evaluation of acoustic telemetry grids for determining aquatic animal movement and survival. Methods in Ecology and Evolution, 9(6), 1489–1502. https://doi.org/10.1111/2041-210X.12996
Lacroix, G. L., McCurdy, P., and Knox, D. 2004. Migration of Atlantic Salmon Postsmolts in Relation to Habitat Use in a Coastal System. Transactions of the American Fisheries Society, 133(6), 1455–1471. https://doi.org/10.1577/T03-032.1
Le Bris, A., Fisher, J. A. D., Murphy, H. M., Galbraith, P. S., Castonguay, M., Loher, T., and Robert, D. 2018. Migration patterns and putative spawning habitats of Atlantic halibut (Hippoglossus hippoglossus) in the Gulf of St. Lawrence revealed by geolocation of pop-up satellite archival tags. ICES Journal of Marine Science, 75(1), 135–147. https://doi.org/10.1093/icesjms/fsx098
Lembke, C., Lowerre-Barbieri, S., Mann, D., and Taylor, J. C. 2018. Using Three Acoustic Technologies on Underwater Gliders to Survey Fish. Marine Technology Society Journal, 52(6), 39–52. https://doi.org/10.4031/MTSJ.52.6.1
Liu, C., Bank, C., Kersula, M., Cowles, G. W., Zemeckis, D. R., Cadrin, S. X., and McGuire, C. 2019. Movements of Atlantic halibut in the Gulf of Maine based on geolocation. ICES Journal of Marine Science, 76(7), 2020–2032. https://doi.org/10.1093/icesjms/fsz169
Loher, T., Webster, R. A., and Carlile, D. 2017. A test of the detection range of acoustic transmitters and receivers deployed in deep waters of Southeast Alaska, USA. Animal Biotelemetry, 5(1), 27. https://doi.org/10.1186/s40317-017-0142-y
Long, M., Jordaan, A., and Castro-Santos, T. 2023. Environmental factors influencing detection efficiency of an acoustic telemetry array and consequences for data interpretation. Animal Biotelemetry, 11(1), 18. https://doi.org/10.1186/s40317-023-00317-2
Mäkinen, T. S., Niemelä, E., Moen, K., and Lindström, R. 2000. Behaviour of gill-net and rod-captured Atlantic salmon (Salmo salar L.) during upstream migration and following radio tagging. Fisheries Research, 45(2), 117–127. https://doi.org/10.1016/S0165-7836(99)00107-1
Martín, G., Espinoza, M., Heupel, M., and Simpfendorfer, C. A. 2020. Estimating marine protected area network benefits for reef sharks. Journal of Applied Ecology, 57(10), 1969–1980. https://doi.org/10.1111/1365-2664.13706
Masmitja, I., Navarro, J., Gomariz, S., Aguzzi, J., Kieft, B., O’Reilly, T., Katija, K., Bouvet, P. J., Fannjiang, C., Vigo, M., Puig, P., Alcocer, A., Vallicrosa, G., Palomeras, N., Carreras, M., del Rio, J., and Company, J. B. 2020. Mobile robotic platforms for the acoustic tracking of deep-sea demersal fishery resources. Science Robotics, 5(48), eabc3701. https://doi.org/10.1126/scirobotics.abc3701
Matley, J. K., Klinard, N. V., Martins, A. P. B., Aarestrup, K., Aspillaga, E., Cooke, S. J., Cowley, P. D., Heupel, M. R., Lowe, C. G., Lowerre-Barbieri, S. K., Mitamura, H., Moore, J.-S., Simpfendorfer, C. A., Stokesbury, M. J. W., Taylor, M. D., Thorstad, E. B., Vandergoot, C. S., and Fisk, A. T. 2022. Global trends in aquatic animal tracking with acoustic telemetry. Trends in Ecology and Evolution, 37(1), 79–94. https://doi.org/10.1016/j.tree.2021.09.001
Neilson, J. D., Waiwood, K. G., and Smith, S. J. 1989. Survival of Atlantic Halibut (Hippoglossus hippoglossus) Caught by Longline and Otter Trawl Gear. Canadian Journal of Fisheries and Aquatic Sciences, 46(5), 887–897. https://doi.org/10.1139/f89-114
O’Brien, M. H. P., and Secor, D. H. 2021. Influence of thermal stratification and storms on acoustic telemetry detection efficiency: A year-long test in the US Southern Mid-Atlantic Bight. Animal Biotelemetry, 9(1), 8. https://doi.org/10.1186/s40317-021-00233-3
O’Dor, R. K., and M. J. W. Stokesbury. 2009. The Ocean Tracking Network – adding marine animal movements to the Global Ocean Observing System. In J.L. Nielsen et al. (eds.), Tagging and Tracking of Marine Animals with Electronic Devices, Reviews: Methods and Technologies in Fish Biology and Fisheries.
Oliver, M. J., Breece, M. W., Fox, D. A., Haulsee, D. E., Kohut, J. T., Manderson, J., and Savoy, T. 2013. Shrinking the Haystack: Using an AUV in an Integrated Ocean Observatory to Map Atlantic Sturgeon in the Coastal Ocean. Fisheries, 38(5), 210–216. https://doi.org/10.1080/03632415.2013.782861
Pante, E. and Simon-Bouhet, B. 2013. marmap: A Package for Importing, Plotting and Analyzing Bathymetric and Topographic Data in R. PLoS ONE 8(9): e73051. https://doi.org/10.1371/journal.pone.0073051
R Core Team. 2024. R: A language and environment for statistical computing. [Computer software]. R Foundation for Statistical Computing. https://www.R-project.org/
Ransier, K. T., Gatti, P., Le Bris, A., den Heyer, C. E., Claireaux, G., Wringe, B., and Fisher, J. A. D. 2024. Electronic tags reveal high migratory diversity within the largest Atlantic halibut (Hippoglossus hippoglossus) stock. Canadian Journal of Fisheries and Aquatic Sciences, 81(7), 828–846. https://doi.org/10.1139/cjfas-2023-0282
ReefMaster Software Ltd. 2021. ReefMaster [Computer software]. https://reefmaster.com.au/
Rutherford, R. J., and Breeze, H. 2002. The Gully Ecosystem (Canadian Manuscript Report of Fisheries and Aquatic Sciences, 2615; p. 29). Fisheries and Oceans Canada.
Sanderson, B. G., Karsten, R. H., and Hasselman, D. J. 2023. Towards Estimating Probability of Fish–Turbine Encounter: Using Drifters Equipped with Acoustic Tags to Verify the Efficacy of an Array of Acoustic Receivers. Journal of Marine Science and Engineering, 11(8), Article 8. https://doi.org/10.3390/jmse11081592
Secor, D. H. 1999. Specifying divergent migrations in the concept of stock: The contingent hypothesis. Fisheries Research, 43(1), 13–34. https://doi.org/10.1016/S0165-7836(99)00064-8
Stokesbury, M. J. W., Logan-Chesney, L. M., McLean, M. F., Buhariwalla, C. F., Redden, A. M., Beardsall, J. W., Broome, J. E., and Dadswell, M. J. 2016. Atlantic Sturgeon Spatial and Temporal Distribution in Minas Passage, Nova Scotia, Canada, a Region of Future Tidal Energy Extraction. PLOS ONE, 11(7), e0158387. https://doi.org/10.1371/journal.pone.0158387
Trumble, R. J. 1993. Atlantic Halibut (Hippoglossus hippoglossus) and Pacific Halibut (H. stenolepis) and Their North American Fisheries. NRC Research Press.
Trzcinski, M., and Bowen, W. 2016. The recovery of Atlantic halibut: A large, long-lived, and exploited marine predator. ICES Journal of Marine Science: Journal Du Conseil, 73, fsv266. https://doi.org/10.1093/icesjms/fsv266
Whitlock, R., Schallert, R. J., Stokesbury, M. J. W., Castleton, M., and Block, B. A. 2022. Informing Management of Atlantic Bluefin Tuna Using Telemetry Data. Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.771547
Zemeckis, D. R., Dean, M. J., DeAngelis, A. I., Van Parijs, S. M., Hoffman, W. S., Baumgartner, M. F., Hatch, L. T., Cadrin, S. X., and McGuire, C. H. 2019. Identifying the distribution of Atlantic cod spawning using multiple fixed and glider-mounted acoustic technologies. ICES Journal of Marine Science, 76(6), 1610–1625. https://doi.org/10.1093/icesjms/fsz064
Zemeckis, D. R., Hoffman, W. S., Dean, M. J., Armstrong, M. P., and Cadrin, S. X. 2014. Spawning site fidelity by Atlantic cod (Gadus morhua) in the Gulf of Maine: Implications for population structure and rebuilding. ICES Journal of Marine Science, 71(6), 1356–1365. https://doi.org/10.1093/icesjms/fsu117
Citation: Bateman, E.M.T., Harper, D.L. , Byrne, T., Comeau, A., den Heyer, C.E., and Stokesbury, M.J.W. 2025. Movement and behaviour of Atlantic Halibut (
Hippoglossus hippoglossus) in a juvenile hotspot detected by a passive moored acoustic receiver grid and active wave gliders.
J. Northw. Atl. Fish. Sci.,
56: 11–29. https://doi.org/10.2960/J.v56.m753